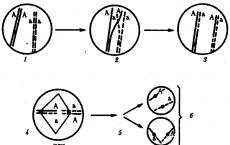
Is the theory of linked inheritance confirmed cytologically? Linked inheritance of genes. Chromosomal theory of heredity T. Morgan. Review questions and assignments
Linked inheritance - inheritance of traits whose genes are localized on the same chromosome. The strength of linkage between genes depends on the distance between them: the farther the genes are located from each other, the higher the frequency of crossing over and vice versa. Along with traits that are inherited independently, there must also be those that are inherited linked to each other, since they are determined by genes located on the same chromosome. Such genes form clutch group. The number of linkage groups in organisms of a certain species is equal to the number of chromosomes in the haploid set (for example, in Drosophila 1 pair = 4, in humans 1 pair = 23).
Full grip- a kind of linked inheritance, in which the genes of the analyzed traits are located so close to each other that crossing over between them becomes impossible.
Incomplete clutch- a type of linked inheritance, in which the genes of the analyzed traits are located at a certain distance from each other, which makes crossing over between them possible.
(Crossover gametes- gametes, in the process of formation of which there was a crossing-over. As a rule, crossover gametes make up a small part of the total number of gametes.
Crossing over- exchange of sections of homologous chromosomes in the process of cell division, mainly in the prophase of the first meiotic division, sometimes in mitosis. The experiments of T. Morgan, K. Bridges and A. Sturtevant showed that there is no absolutely complete linkage of genes, in which genes would always be transmitted together. The probability that two genes located on the same chromosome will not diverge during meiosis ranges from 1-0.5. In nature, incomplete linkage predominates due to the crossing of homologous chromosomes and recombination of genes. The cytological pattern of crossing over was first described by the Danish scientist F. Janssens.
Crossing over occurs only when the genes are in a heterozygous state (AB / av). If the genes are in a homozygous state (AB / AB or aB / aB), the exchange of identical regions does not give new combinations of genes in gametes and in a generation. The frequency (percentage) of crossover between genes depends on the distance between them: the further they are located from each other, the more often crossing over occurs. T. Morgan proposed to measure the distance between genes by crossing over as a percentage, according to the formula:
N1/N2 X 100 = % Crossover,
where N1 is the total number of individuals in F;
N2 is the total number of crossover individuals.
The segment of the chromosome, on which 1% of crossing over takes place, is equal to one morganide (a conditional measure of the distance between genes). The frequency of crossing over is used to determine the relative position of genes and the distance between them. To build a human genetic map, new technologies are used, in addition, cytogenetic maps of chromosomes have been built.
There are several types of crossing over: double, multiple (complex), irregular, uneven.
Crossing over leads to a new combination of genes, causing a change in the phenotype. In addition, along with mutations, it is an important factor in the evolution of organisms.)
The result of T. Morgan's research was his creation of the chromosome theory of heredity:
· genes are located on chromosomes; different chromosomes contain an unequal number of genes; the set of genes for each of the nonhomologous chromosomes is unique;
· each gene has a specific location (locus) on the chromosome; allelic genes are located in identical loci of homologous chromosomes;
· genes are located on chromosomes in a certain linear sequence;
· genes located on the same chromosome are inherited together, forming a linkage group; the number of linkage groups is equal to the haploid set of chromosomes and is constant for each type of organism;
· gene linkage can be broken during the process of crossing over, which leads to the formation of recombinant chromosomes; the frequency of crossing over depends on the distance between genes: the greater the distance, the greater the value of crossing over;
· each species has a set of chromosomes characteristic only for it - a karyotype.
Inheritance of sex and sex-linked traits. Sex chromosomes and their role in sex determination. Sex inheritance. The sex of an individual is a complex trait, formed both by the action of genes and by the conditions of development. In humans, one of the 23 pairs of chromosomes is the sex chromosomes, denoted as X and Y. Women are homogametic sex, i.e. They have two X chromosomes, one from their mother and one from their father. Males are heterogametic, have one X-one Y-chromosome, and X is transmitted from the mother, and Y from the father. Note that the heterogametic sex is not always necessarily male; for example, in birds, these are females, while males are homogametic. There are other mechanisms of sex determination. So, in a number of insects, the Y chromosome is absent. In this case, one of the sexes develops in the presence of two X chromosomes, and the other - in the presence of one X chromosome. In some insects, sex is determined by the ratio of the number of autosomes and sex chromosomes. In a number of animals, the so-called. redefinition of sex, when, depending on environmental factors, the zygote develops either into a female or into a male. The development of sex in plants has as diverse genetic mechanisms as in animals.
Traits linked to the X chromosome. If the gene is located on the sex chromosome (it is called sex-linked), then its expression in descendants follows rules other than for autosomal genes. Consider genes located on the X chromosome. A daughter inherits two X chromosomes, one from her mother and one from her father. The son has only one X-chromosome - from the mother; from his father he receives a Y-chromosome. Therefore, the father passes on the genes on his X chromosome only to his daughter, while the son cannot receive them. Since the X chromosome is more "rich" in genes compared to the Y chromosome, in this sense, the daughter is genetically more similar to the father than the son; the son is more like his mother than his father.
One of the historically best known sex-linked traits in humans is hemophilia, resulting in severe bleeding from minor cuts and extensive bruising from bruises. It is caused by a recessive defective allele 0, which blocks the synthesis of a protein necessary for blood clotting. The gene for this protein is located on the X chromosome. A heterozygous woman +0 (+ means the normal active allele, dominant in relation to the hemophilia 0 allele) does not get hemophilia, and her daughters, too, if the father does not have this pathology. However, her son can get the 0 allele, and then he develops hemophilia. Recessive diseases caused by X-chromosome genes affect women much less frequently than men, since they have the disease only when they are homozygous - the presence of a recessive allele in each of the two homologous X chromosomes; males get sick whenever their single X chromosome carries the defective allele.
Linkage to the Y chromosome.Information about the genes located on the Y chromosome is very scarce. It is assumed that it practically does not carry the genes that determine the synthesis of proteins necessary for the functioning of the cell. But it plays a key role in the development of the male phenotype. The absence of a Y chromosome in the presence of only one X chromosome leads to the so-called. Turner syndrome: the development of a female phenotype with poorly developed primary and secondary sexual characteristics and other abnormalities. There are men with an additional Y-chromosome (XYY); they are tall, aggressive and often have abnormal behavior. In the Y chromosome, several genes have been identified that are responsible for regulating the synthesis of specific enzymes and hormones, and violations in them lead to pathologies of sexual development. There are a number of morphological traits that are believed to be determined by genes on the Y chromosome; among them is the development of ear hair. Such signs are transmitted only through the male line: from father to son.
genetic sex determination, determined by the set of sex chromosomes, supports equal reproduction of females and males. Indeed, female eggs contain only the X chromosome, since women have the XX genotype on the sex chromosomes. The genotype of men is XY, and therefore the birth of a girl or a boy in each case is determined by whether the sperm carries the X or Y chromosome. Since in the process of meiosis the chromosomes have an equal chance of getting into the gamete, then half of the gametes produced by male individuals contain the X-chromosome, and half the Y-chromosome. Therefore, half of the offspring are expected to be of one sex and half of the other.
It should be emphasized that it is impossible to predict the birth of a boy or a girl in advance, since it is impossible to predict which male germ cell will participate in the fertilization of the egg: the one carrying the X- or Y-chromosome. Therefore, the presence of more or less boys in the family is a matter of chance.
Analysis of the phenomena of linked inheritance, crossing over, comparison of genetic and cytological maps allow us to formulate the main provisions of the chromosome theory of heredity:
Genes are located on chromosomes. Moreover, different chromosomes contain an unequal number of genes. In addition, the set of genes for each of the non-homologous chromosomes is unique.
Allelic genes occupy the same loci on homologous chromosomes.
Genes are located on the chromosome in a linear sequence.
The genes of one chromosome form a linkage group, that is, they are inherited predominantly linked (jointly), due to which the linked inheritance of some traits occurs. The number of linkage groups is equal to the haploid number of chromosomes of a given species (in the homogametic sex) or more by 1 (in the heterogametic sex).
Linkage is broken as a result of crossing over, the frequency of which is directly proportional to the distance between the genes in the chromosome (therefore, the linkage strength is inversely related to the distance between the genes).
Each biological species is characterized by a certain set of chromosomes - karyotype.
Linked inheritance
Independent combination of features ( Mendel's third law) is carried out under the condition that the genes that determine these traits are in different pairs of homologous chromosomes. Therefore, in each organism, the number of genes that can independently combine in meiosis is limited by the number of chromosomes. However, in an organism, the number of genes significantly exceeds the number of chromosomes. For example, before the era of molecular biology, more than 500 genes were studied in corn, more than 1 thousand in the Drosophila fly, and about 2 thousand genes in humans, while they have 10, 4 and 23 pairs of chromosomes, respectively. The fact that the number of genes in higher organisms is several thousand was already clear to W. Setton at the beginning of the 20th century. This gave reason to assume that many genes are localized in each chromosome. Genes located on the same chromosome form a linkage group and are inherited together.
Joint inheritance of genes T. Morgan proposed to name linked inheritance. The number of linkage groups corresponds to the haploid number of chromosomes, since the linkage group consists of two homologous chromosomes in which the same genes are localized. (In individuals of the heterogametic sex, for example, male mammals, there are actually one more linkage groups, since the X and Y chromosomes contain different genes and represent two different linkage groups. Thus, women have 23 linkage groups, and in men - 24).
The mode of inheritance of linked genes differs from the inheritance of genes located in different pairs of homologous chromosomes. So, if, with independent combination, a diheterozygous individual forms four types of gametes (AB, Ab, aB and ab) in equal quantities, then with linked inheritance (in the absence of crossing over), the same diheterozygote forms only two types of gametes: (AB and ab) also in equal amounts. The latter repeat the combination of genes in the parent's chromosome.
It was found, however, that in addition to ordinary (non-crossover) gametes, other (crossover) gametes also arise with new gene combinations - Ab and aB, which differ from the combinations of genes in the parent's chromosomes. The reason for the formation of such gametes is the exchange of sections of homologous chromosomes, or crossing over.
Crossing over occurs in prophase I of meiosis during conjugations homologous chromosomes. At this time, parts of two chromosomes can cross over and exchange their parts. As a result, qualitatively new chromosomes arise, containing sections (genes) of both maternal and paternal chromosomes. Individuals that are obtained from such gametes with a new combination of alleles are called crossing-over or recombinant.
The frequency (percentage) of crossover between two genes located on the same chromosome is proportional to the distance between them. Crossing over between two genes occurs less frequently the closer they are to each other. As the distance between genes increases, the likelihood that crossing over will separate them on two different homologous chromosomes increases more and more.
The distance between genes characterizes the strength of their linkage. There are genes with a high percentage of linkage and those where linkage is almost not detected. However, with linked inheritance, the maximum crossover frequency does not exceed 50%. If it is higher, then there is a free combination between pairs of alleles, indistinguishable from independent inheritance.
The biological significance of crossing over is extremely high, since the genetic recombination allows you to create new, previously non-existing combinations of genes and thereby increase hereditary variability, which gives ample opportunities for adaptation of the organism in various environmental conditions. A person specially conducts hybridization in order to obtain the necessary combinations for use in breeding work.
Crossing over. This process occurs in prophase I of meiosis at a time when homologous chromosomes are closely brought together as a result of conjugation and form bivalents. During crossing over, the corresponding sections are exchanged between mutually intertwining chromatids of homologous chromosomes (Fig. 3.72). This process ensures the recombination of paternal and maternal alleles of genes in each linkage group. In different precursors of gametes, crossing over occurs in different parts of the chromosomes, resulting in a wide variety of combinations of parental alleles in the chromosomes.
Rice. 3.72. Crossing over as a source of genetic diversity of gametes:
I - fertilization of parental gametes a and b c zygote formation in;II - gametogenesis in an organism developed from a zygote in;G- crossing over between homologues in prophase I; d - cells formed after the 1st meiotic division; e, w - cells formed after the 2nd division of meiosis ( e - non-crossover gametes with original parental chromosomes; well - crossover gametes with recombination of hereditary material in homologous chromosomes)
It is clear that crossing over as a mechanism of recombination is effective only when the corresponding genes of the paternal and maternal chromosomes are represented by different alleles. Absolutely identical linkage groups during crossing over do not give new combinations of alleles.
Crossing over occurs not only in the precursors of germ cells during meiosis. It is also observed in somatic cells during mitosis. Somatic crossing-over has been described in Drosophila and in some mold species. It is carried out during mitosis between homologous chromosomes, but its frequency is 10,000 times less than the frequency of meiotic crossing over, from the mechanism of which it does not differ. As a result of mitotic crossing over, clones of somatic cells appear that differ in the content of individual gene alleles in them. If in the genotype of the zygote this gene is represented by two different alleles, then as a result of somatic crossing over, cells with the same either paternal or maternal alleles of this gene may appear (Fig. 3.73).

Rice. 3.73. Crossing over in somatic cells:
1 - a somatic cell, in the homologous chromosomes of which gene A is represented by two different alleles (A and a); 2 - crossing over; 3 - the result of the exchange of the corresponding sections between homologous chromosomes; 4 - location of homologues in the equatorial plane of the fission spindle in the metaphase of mitosis (two variants); 5 - formation of daughter cells; 6 - the formation of cells heterozygous for gene A, similar to the mother cell in terms of a set of alleles (Aa); 7 - the formation of cells homozygous for gene A, which differ from the mother cell in the set of alleles (AA or aa)
In the middle of the 19th century, when G. Mendel carried out his experiments and formulated patterns of universal and fundamental importance for the development of genetics and biology in general, scientific knowledge was still not enough to understand the mechanisms of inheritance. That is why for many years the work of Mendel was unclaimed. However, by the beginning of the 20th century. the situation in biology has changed radically.
Mitosis and meiosis were discovered, Mendel's laws were rediscovered. Independently of each other, researchers in Germany and the United States proposed that hereditary factors are located on the chromosomes. In 1906, R. Pennet first described the violation of the Mendelian law of independent inheritance of two traits. When staging a classic dihybrid cross of sweet pea plants that differ in flower color and pollen shape, Pennet did not get the expected splitting of 9:3:3:1 in the second generation. F2 hybrids had only parental phenotypes in a ratio of 3:1, i.e., no redistribution of traits occurred.
Gradually, more and more such exceptions accumulated, which did not obey the law of independent inheritance. The question arose, how exactly are genes located on chromosomes? After all, the number of traits, and therefore the number of genes in each organism is much greater than the number of chromosomes. This means that each chromosome contains many genes responsible for different traits. How are genes located on the same chromosome inherited?
■ The work of T. Morgan. These questions were answered by a group of American scientists headed by Thomas Hunt Morgan (1866-1945). Working on a very convenient genetic object - the Drosophila fruit fly, they did a great job of studying the inheritance of genes.
Scientists have found that genes located on the same chromosome are inherited together, i.e. linked. This phenomenon has been named morgan law, or the law of linked inheritance . Groups of genes located on the same chromosome are called clutch group. Since identical genes are located in homologous chromosomes, the number of linkage groups is equal to the number of pairs of chromosomes, i.e., the haploid number of chromosomes. A person has 23 pairs of chromosomes and, therefore, 23 linkage groups, a dog has 39 pairs of chromosomes and 39 linkage groups, a pea has 7 pairs of chromosomes and 7 linkage groups, etc. It should be noted that Mendel was surprisingly lucky when setting up di-hybrid crosses : genes responsible for different traits (color and shape of peas) were located on different chromosomes. It could have been otherwise, and then the pattern of independent splitting would not have been discovered.
The result of the work of T. Morgan's group was the creation in 1911 of chromosome theory of heredity.
Consider the main provisions of the modern chromosome theory of heredity.
The unit of heredity is a gene, which is a segment of a chromosome.
Genes are located in chromosomes in strictly defined places (locuses), and allelic genes (responsible for the development of one trait) are located in the same loci of homologous chromosomes.
Genes are located on chromosomes in a linear order, that is, one after another.
■Clutch failure. However, in some crosses, when analyzing the inheritance of genes located on the same chromosome, a linkage disorder was found. It turned out that sometimes paired homologous chromosomes can exchange identical homologous regions with each other. In order for this to happen, the chromosomes must be located in close proximity to each other. This temporary pairwise approach of homologous chromosomes is called conjugation. In this case, the chromosomes can exchange loci located opposite each other containing the same genes. This phenomenon is called crossing over.
Recall the division of meiosis, during which germ cells are formed. In the prophase of the first meiotic division, during the formation of a bivalent (tetrad), when duplicated homologous chromosomes stand parallel to each other, a similar exchange can occur. Such an event leads to a recombination of genetic material, increases the diversity of descendants, i.e., increases hereditary variability and, therefore, plays an important role in evolution.
■ Genetic maps. The phenomenon of the exchange of allelic genes between homologous chromosomes helped scientists to determine the location of each gene on the chromosome, that is, to build genetic maps. The genetic map of a chromosome is a diagram of the mutual arrangement of genes located on the same chromosome, i.e., in the same linkage group. The construction of such maps is of great interest both for fundamental research and for solving a variety of practical problems. For example, genetic maps of human chromosomes are very important for diagnosing a number of severe hereditary diseases.
At present, simple genetic maps are being replaced by molecular genetic maps, which contain information about the nucleotide sequences of genes.
Questions for self-control
1. What is linked inheritance?
2. What are gene linkage groups?
3. What is the cause of gene linkage disorder?
4. What is the biological significance of the exchange of allelic genes between homologous chromosomes?
5. Is the theory of linked inheritance confirmed cytologically?
According to G. Mendel, genes are inherited independently and are located on different chromosomes. Diheterozygote AaBb in the form of a cytological formula can be represented as follows:
During meiosis, it forms 4 types of gametes: AB, Ab, aB, ab.
In 1911–1912 American geneticist T. Morgan proved that genes located on the same chromosome can be inherited linked (together). This is due to the fact that there are many more genes than chromosomes. Co-inheritance of genes limiting their free combination, T. Morgan called gene linkage, or linked inheritance. In the case when the genes are inherited linked, the genotype of the AaBb diheterozygote using the cytological formula can be represented as follows:
During meiosis, if there is complete linkage of genes, it forms 2 types of gametes - AB, ab. Such gametes are called non-crossover.
Research by T. Morgan and his students showed that genes are regularly exchanged in a homologous pair of chromosomes. The process of exchanging identical sections of homologous chromosomes with the genes located in them is called chromosome decussation, or crossing over. Crossing over provides new combinations of genes located on homologous chromosomes . Therefore, due to the phenomenon of crossing over, in addition to non-crossover, there are also crossover gametes - Ab, aB.
T. Morgan's Law is formulated as follows: genes located on the same chromosome are inherited linked (together), and the strength of the link depends on the distance between the genes.
The main provisions of T. Morgan's chromosome theory:
1) genes are localized on chromosomes, and different chromosomes contain an unequal number of genes. The set of genes in each of the non-homologous chromosomes is unique;
2) genes are located on the chromosome in a certain linear sequence;
3) genes form linkage groups and can be inherited linked. The number of linkage groups corresponds to the haploid set of chromosomes (a person has 23 linkage groups);
4) between homologous chromosomes, a crossing over (crossover) and an exchange of allelic genes is possible, and the crossover occurs when there is a distance between the genes;
5) the distance between genes in the chromosome is proportional to the frequency of crossing and is expressed as a percentage of crossing over.
The percentage of crossing over between genes is calculated by the formula:
X= ––––
a is the number of crossover individuals of one class; b is the number of crossover individuals of another class;
n is the total number of individuals; Х – % of crossing over between genes (distance between genes in % of crossing over).
The distance between genes is measured in special units - morganides. 1 morganide (1 M) = 1% crossing over.
In genetic studies, a fruit fly was used, which has a karyotype consisting of 8 chromosomes, has fertility, and many alternative traits. T. Morgan considered the situation when complete linkage of genes and violation of linkage of genes (crossing over) occur in an individual at the same time. At the first stage, homozygous individuals (P ♀AABB x ♂aabb) were taken for crossing. As a result, in the first generation, all individuals were diheterozygous (AaBb).
Gene Trait
A gray body color
and dark body color
In long wings
b rudimentary wings.
Full linkage of genes
P ♀ ══x ♂ ══
G AB, ab ab - male gametes
(Non-crossover female gametes)
F 1 ══ ; ══
With complete linkage of genes, a 1:1 split is observed. In total, 83% of individuals were obtained that inherited the phenotype of their parents, since they were formed from non-crossover gametes. Among them, individuals with a gray body, normal wings accounted for 41.5% and individuals with a black body, rudimentary wings also accounted for 41.5%.
Gene linkage disorder
P ♀ ══ x ♂ ══
G Ab, aB ab - male gametes
(Crossover female gametes)
F 1 ══; ══
With incomplete linkage of genes, 1:1 splitting is also observed. In total, 17% of individuals with recombinant phenotypes were obtained, since they were formed from crossover gametes. Individuals with a new combination of traits are called recombinants. Among the recombinants, flies with a gray body, rudimentary wings accounted for 8.5%, and with a black body, normal wings also accounted for 8.5%.
Question 1. What is linked inheritance?
Linked inheritance is the joint inheritance of genes located on the same chromosome (i.e., in one DNA molecule). For example, in sweet peas, the genes that determine the color of flowers and the shape of pollen are arranged in this way. They are inherited in a linked manner, therefore, when crossing hybrids of the second generation, parental phenotypes are formed in a ratio of 3: 1, and the splitting 9: 3: 3: 1, characteristic of dihybrid crossing with independent inheritance, does not appear.
Question 2. What are gene linkage groups?
A linkage group is a collection of genes located on the same chromosome. Since the homologous chromosomes contain the same genes, the number of linkage groups is equal to the haploid number of chromosomes (23 in humans, 7 in peas, 4 in Drosophila).
Question 3. What is the cause of gene linkage disorder?
The reason for the disruption of gene linkage is the exchange of sections of homologous chromosomes in prophase I of meiotic division. Recall that at this stage, paired chromosomes are conjugated, forming the so-called bivalent bands. The formation of bivalents can lead to the crossover of chromosomes, which creates the possibility of the exchange of homologous DNA regions. If this happens, then the linkage groups change their content (they contain other alleles of the same genes) and individuals with a phenotype that differs from the parent ones may appear in the offspring.
Question 4. What is the biological significance of the exchange of allelic genes between homologous chromosomes?material from the site
The crossing of chromosomes (otherwise - crossing over) leads to the recombination of genetic material and the formation of new combinations of alleles of genes from the linkage group. At the same time, the diversity of descendants increases, i.e., hereditary variability increases, which is of great evolutionary importance. Indeed, if, for example, in Drosophila, the genes that determine the color of the body and the length of the wings are on the same chromosome, then, crossing pure lines of gray flies with normal wings and black flies with shortened wings, in the absence of crossing over, we will never get other phenotypes. The existence of a crossover of chromosomes allows the appearance (in a few percent of cases) of gray flies with short wings and black flies with normal wings.
Question 5. Has the theory of linked inheritance been confirmed cytologically?
The theory of linked inheritance by Thomas Hunt Morgan (1866-1945) is confirmed by cytological observations. It was shown that during division, chromosomes completely diverge to different poles of the cell. Consequently, genes located on the same chromosome during meiosis fall into one gamete, that is, they are really inherited in a linked fashion.
Didn't find what you were looking for? Use the search