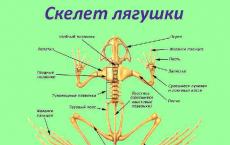
Frog skull structure. The structure of amphibians. Structural features of the frog

 Features of the frog skeleton The spine consists of 1 cervical, 7 trunk, 1 sacral vertebrae and the caudal section (urostyle) There are no ribs or rib cage
Features of the frog skeleton The spine consists of 1 cervical, 7 trunk, 1 sacral vertebrae and the caudal section (urostyle) There are no ribs or rib cage
 Features of amphibians: greater differentiation of the spinal column into sections and a stronger connection of the vertebral bodies with each other; the appearance of paired terrestrial limbs; increasing complexity of the structure and greater strength of the limb girdles; strong reduction of metameric muscles and its replacement by a fairly powerful complex complex of muscles; the appearance of eyelids (protecting the eyes from mechanical damage, preventing the cornea from drying out, etc.); formation of the middle ear cavity with the eardrum and the auditory ossicle - the stapes (ensuring the possibility of hearing in the air). the disappearance of gills and the development of lungs, larynx and choanae, creating the possibility of air breathing; the emergence of two circles of blood circulation; greater differentiation of the digestive system (high energy costs when maintaining the body in the air); general lengthening of the hind limbs, isolation of an additional lever in them
Features of amphibians: greater differentiation of the spinal column into sections and a stronger connection of the vertebral bodies with each other; the appearance of paired terrestrial limbs; increasing complexity of the structure and greater strength of the limb girdles; strong reduction of metameric muscles and its replacement by a fairly powerful complex complex of muscles; the appearance of eyelids (protecting the eyes from mechanical damage, preventing the cornea from drying out, etc.); formation of the middle ear cavity with the eardrum and the auditory ossicle - the stapes (ensuring the possibility of hearing in the air). the disappearance of gills and the development of lungs, larynx and choanae, creating the possibility of air breathing; the emergence of two circles of blood circulation; greater differentiation of the digestive system (high energy costs when maintaining the body in the air); general lengthening of the hind limbs, isolation of an additional lever in them
 “Aquatic” structural features are manifested in a number of features: the relatively weak development of skeletal ossifications, the abundance of mucous glands in the skin (the mucus covering the skin reduces friction when moving in water, prevents the penetration of bacteria and fungi into the skin, etc.), preservation of the tail, often flattened laterally and bordered by a leathery fold (newts and other tailed amphibians), great similarity of the genitourinary system with most groups of fish, external fertilization characteristic of the vast majority of amphibian species, etc.
“Aquatic” structural features are manifested in a number of features: the relatively weak development of skeletal ossifications, the abundance of mucous glands in the skin (the mucus covering the skin reduces friction when moving in water, prevents the penetration of bacteria and fungi into the skin, etc.), preservation of the tail, often flattened laterally and bordered by a leathery fold (newts and other tailed amphibians), great similarity of the genitourinary system with most groups of fish, external fertilization characteristic of the vast majority of amphibian species, etc.
 With a relatively small surface area of the lungs of amphibians, fairly powerful additional respiratory organs are required. Such an organ becomes the skin that is always moist (due to the abundance of mucous glands), easily permeable to moisture and gases, and partly the mucous membrane of the oral cavity. In an active pond frog, the lungs absorb up to 50% of the oxygen consumed by the body and release only about 14% of carbon dioxide; through skin respiration, up to 50% of oxygen is absorbed and up to 86% of carbon dioxide is released. In the grass frog, which leads a more terrestrial life, pulmonary respiration takes in up to 67% of oxygen and releases up to 26% of carbon dioxide, and through skin respiration, 33% of oxygen is absorbed and 74% of carbon dioxide is released. With an increase in the level of metabolism (increased general activity and all metabolic processes with increasing environmental temperature), the specific role of the lungs in providing the body with oxygen increases markedly. A decrease in environmental temperature causes a decrease in the level of metabolism. At the same time, skin respiration almost completely ensures both the saturation of the body with oxygen and the release of carbon dioxide, and the relative importance of the lungs in breathing sharply decreases.
With a relatively small surface area of the lungs of amphibians, fairly powerful additional respiratory organs are required. Such an organ becomes the skin that is always moist (due to the abundance of mucous glands), easily permeable to moisture and gases, and partly the mucous membrane of the oral cavity. In an active pond frog, the lungs absorb up to 50% of the oxygen consumed by the body and release only about 14% of carbon dioxide; through skin respiration, up to 50% of oxygen is absorbed and up to 86% of carbon dioxide is released. In the grass frog, which leads a more terrestrial life, pulmonary respiration takes in up to 67% of oxygen and releases up to 26% of carbon dioxide, and through skin respiration, 33% of oxygen is absorbed and 74% of carbon dioxide is released. With an increase in the level of metabolism (increased general activity and all metabolic processes with increasing environmental temperature), the specific role of the lungs in providing the body with oxygen increases markedly. A decrease in environmental temperature causes a decrease in the level of metabolism. At the same time, skin respiration almost completely ensures both the saturation of the body with oxygen and the release of carbon dioxide, and the relative importance of the lungs in breathing sharply decreases.
 This duality in the nature of breathing is explained not only by the insufficient development of the surface of the lungs and the imperfection of pulmonary ventilation (“swallowing” of air in the absence of a chest); it is necessary for the amphibian lifestyle of representatives of this class. It is this duality of the respiratory organs that provides amphibians with the opportunity to stay in water for a long time (up to wintering at the bottom of the reservoir of many species of anurans, when, with a sharp decrease in the level of metabolism, skin respiration completely meets all the body’s needs for oxygen and the release of carbon dioxide).
This duality in the nature of breathing is explained not only by the insufficient development of the surface of the lungs and the imperfection of pulmonary ventilation (“swallowing” of air in the absence of a chest); it is necessary for the amphibian lifestyle of representatives of this class. It is this duality of the respiratory organs that provides amphibians with the opportunity to stay in water for a long time (up to wintering at the bottom of the reservoir of many species of anurans, when, with a sharp decrease in the level of metabolism, skin respiration completely meets all the body’s needs for oxygen and the release of carbon dioxide).

 Internal structure of the frog Digestive system: salivary glands, cloaca Respiratory system: paired lungs, breathing through the skin Circulatory system: 3-chambered heart, 2 circles of blood circulation Nervous system: the forebrain is well developed, the cerebellum is underdeveloped Sensory organs: the middle ear appears, the sense of smell is well developed
Internal structure of the frog Digestive system: salivary glands, cloaca Respiratory system: paired lungs, breathing through the skin Circulatory system: 3-chambered heart, 2 circles of blood circulation Nervous system: the forebrain is well developed, the cerebellum is underdeveloped Sensory organs: the middle ear appears, the sense of smell is well developed

 How a toad catches crawling prey 1. The toad notices the prey in the lateral field of vision of one eye. 2. The toad turns to the prey with the center of the field of vision of this eye. This is how the eye sees most clearly.
How a toad catches crawling prey 1. The toad notices the prey in the lateral field of vision of one eye. 2. The toad turns to the prey with the center of the field of vision of this eye. This is how the eye sees most clearly.
 How a toad catches crawling prey 3. Then it turns and the center of the field of vision of the second eye. 4. Using binocular vision, the toad accurately shoots its sticky tongue at its prey.
How a toad catches crawling prey 3. Then it turns and the center of the field of vision of the second eye. 4. Using binocular vision, the toad accurately shoots its sticky tongue at its prey.
 The frog only catches moving prey with its long sticky tongue. It follows the fly with moving eyes, and its brain calculates the meeting point and commands to the leg muscles.
The frog only catches moving prey with its long sticky tongue. It follows the fly with moving eyes, and its brain calculates the meeting point and commands to the leg muscles.

 The structure of the heart The heart of a fish The heart of a frog Is there a difference in the structure of the hearts of a frog and a fish?
The structure of the heart The heart of a fish The heart of a frog Is there a difference in the structure of the hearts of a frog and a fish?




 Amphibians are dioecious animals that reproduce in water. Females lay eggs, males secrete seminal fluid. Fertilization is external. Development with metamorphosis: the eggs hatch into fish-like tadpoles, which develop into adult amphibians during development. The lifestyle of amphibians changes depending on seasonal changes in living conditions.
Amphibians are dioecious animals that reproduce in water. Females lay eggs, males secrete seminal fluid. Fertilization is external. Development with metamorphosis: the eggs hatch into fish-like tadpoles, which develop into adult amphibians during development. The lifestyle of amphibians changes depending on seasonal changes in living conditions.
 Caring for the Young Most frogs and toads lay their eggs in water and leave them unattended. However, many species of amphibians exhibit interesting forms of caring for their offspring. Thus, the female Surinamese pipa bears eggs in skin cells on her back. The male Darwin's rhinoderma stores the eggs in a special throat pouch. Tree snapping frogs, native to Africa, lay their eggs in a foamy secretion on the leaves of trees located above the water. The hatched tadpoles fall straight into the water. The female bicolor poison dart frog lays eggs in water-filled leaf axils. The larvae emerging from the eggs stick to the back of the male, who carries them to “individual” reservoirs.
Caring for the Young Most frogs and toads lay their eggs in water and leave them unattended. However, many species of amphibians exhibit interesting forms of caring for their offspring. Thus, the female Surinamese pipa bears eggs in skin cells on her back. The male Darwin's rhinoderma stores the eggs in a special throat pouch. Tree snapping frogs, native to Africa, lay their eggs in a foamy secretion on the leaves of trees located above the water. The hatched tadpoles fall straight into the water. The female bicolor poison dart frog lays eggs in water-filled leaf axils. The larvae emerging from the eggs stick to the back of the male, who carries them to “individual” reservoirs.

General remarks
The frog skeleton normally has 80 bones, counting each paired bone as one. The number of otos may vary somewhat due to the sesaado-shaped bones, the degree of ossification of the carpal and tarsal formations and variations in the structure of the spine. For the purpose of detailed study, it is recommended to dissect the skeleton, separating the skeleton of the head, the axial skeleton, the girdles of the fore and hind limbs and the skeleton of the limbs themselves.
The skeleton of an adult grass, or green, frog weighs from 2.5 to 6 g when fresh, which is 9-11% of the weight of the entire corpse. Within an hour, it loses 4% of its original weight due to drying. The finally dried skeleton weighs on average 62-67% of its original weight. There are cyclical changes in the total weight of the skeleton, mainly due to the skeleton of the forelimbs: in the spring it is heavier than in the fall, which is especially noticeable in males. (set for lake and grass frogs). The skeleton of males is generally coarser than that of females.
Organic matter makes up approximately 36% of the total bone weight.
Head skeleton
The formal concept of the head skeleton actually covers formations that are quite heterogeneous in their history: the braincase, the facial skeleton and the hyoid apparatus. However, topographic relationships and close functional connections dictate their joint study.
Upon general examination, what is striking is, firstly, the general shape of the skull: it is relatively flat, more or less rounded at the front. Secondly, the openwork obtained due to the large eye sockets, the window between the jaw and nasal bones and the gap between the quadratojugal and pterygoid bones is interesting. Finally, it is important to note the presence of a significant amount of cartilage. The fact of a greater predominance of cartilage in the skull of anurans compared to the fossil ancestors of amphibians (stegocephalians) is beyond doubt. The main bones corresponding to certain parts of the primary skull (autostoses, or ossa substituentia) in the frog are: the lateral occipital, the anterior auricle, the girdle bone, part of the quadratojugal, the central part of the dental, the middle part of the auditory column and the mid-posterior process of the hyoid apparatus. The integumentary bones (“allostoses”, or ossa investientia) will be: frontoparietal, integumentary sphenoid, nasal, vomer, palatine, maxillary, premaxillary, internasal, pterygoid, squamosal, part of the quadratojugal, part of the tooth and angular.
Rice. 1. General view of the skeleton of a male frog.

Rice. 2. Dismembered skeleton of a female lake frog.



Rice. 3. Lake Frog Skull:
a - top view, 6 - bottom view, c - side view, d - rear view (cartilages are indicated by a dotted line); 1 - premaxillary bone, 2 - jawbone, 3 - nasal, 4 - girdle bone, 5 - frontoparietal, 6 - anterior auricle, 7 - lateral occipital, 8 - pterygoid, 9 - quadratojugal, 10 - squamosal, 11 - vomer, 12 - palatine , 13 - integumentary wedge-shaped, 14 - occipital condyle, 15 - tympanic membrane, 16 - Meckel-mental cartilage, 17 - dental, 18 - angular, 19 - main plate of the hyoid apparatus, 20 - its pterygoid process, 21 - quadrate cartilage, 22 - auditory column, 23 - main horn of the hyoid apparatus, 24 - foramen magnum. Roman numerals indicate the exit points of the corresponding head nerves.
To obtain a complete picture of the primary (primordial) skull, it is most convenient to take a very young frog, but then one has to resort to a rather complex technique of plastic reconstruction. A rougher idea can be obtained by removing the integumentary ossifications from the skull of an adult frog. In the primary skull, the occipital and ear regions, the orbital and nasal regions are distinguished.
In the region of the back of the head and ear (regio occipitalis), two ossifications are formed on each side - the lateral occipital and anterior auricular bones. Each of these bones is separated from the other by a narrow strip of cartilage (synchondrosis prootico-occipitalis). Cartilaginous plates are preserved below and above, connecting the right and left sections. The lower of them can be called the inter-auditory floor (sоlum synoticum), and the upper cartilaginous plate is called the inter-auditory bridge, or roof (tectum sunоticum). The lateral occipital bones (ossa occipitalia lateralia) form the back of the skull and limit the foramen magnum (foramen magnum). In each of these bones, three parts can be distinguished: the medial medial (cranial) (pars cranialis), the lateral labyrinth (parslabyrinthica) and located at the base of the skull (pars basalis). The first of them bears the articular condyle (condulus occipitalis) covered with cartilage. If you look at the occipital condyle from behind, then to the side of it and slightly upward is the articular fossa (fossa condyloidea). Three openings open into it successively: a larger one, located closer to the condyle, the jugular opening (foramen jugulare), and then the upper and lower perilymphatic (for. rerilymphaticum superius et inferius). The IX and X cephalic nerves exit through the first of them. For the exit of the branches of the VIII cerebral nerve, there are two openings on the inner side (from the side of the cranial cavity) of the lateral occipital bone (for. acusticum anterius et posterius). On the whole skeleton, the attachment of the skull to the first vertebra is helped by the longitudinal, unpaired, apical ligament (ligamentum apicis). Its role in tadpoles is performed by the anterior section of the notochord. The occipital joint allows only minor nodding movements. The anterior auricular bone (os prooticum = petrosum) limits the posterior part of the orbit, forming the roof and anterior wall of the auditory capsule. It is distinguished between the cranial part (pars cranialis) and the labyrinthine part (pars labyrinthica), the outer boundary of which is a small ridge on the surface, enclosing the anterior semicircular canal. On the lower part of the anterior auricular bone there are openings for the exit of the V, VI and VII cephalic nerves, as well as for the exit from the cranial cavity of a branch, the internal jugular vein. Posterior to the anterior bone on the cartilaginous skull, a fossa for the oval window (fossa fenestrae ovalis) is visible in the cartilaginous mass, in the depth of which the oval window itself (fenestra ovalis) is located. It leads into the inner cavity of the labyrinth. The posterolateral corner of the skull is occupied by a mass of cartilage homologous to the quadratum of other animals. It consists of a body (corpus quadrati) and three processes. At the posterior end of the body of the quadrate cartilage there is an articular process (processus articularis quadrati). From the anterior end of the body, the main process (pr. basalis quadrati) is directed to the cartilaginous mass of the ear capsule. At the junction of the latter with the cartilaginous skull there is a break in the continuity of the cartilage, reminiscent of a joint. Anterior to the body of the quadrate cartilage is the pterygoid process (pterygoideus quadrati), forming the infraorbital arch (arcus subocularis) and joining the cranium in front of the sphenoid-olfactory bone. Quadrate cartilage also takes part in the formation of the posterior part of the quadratojugal bone. Connected to the quadrate cartilage is the cartilaginous tympanic ring (annulus tympanicus), which separates from it in the larval stage.

Rice. 4. Primary skull of a young (2 cm total length) grass frog. The ear column is removed, and the beginning of ossification is not shown:
1 - pterygoid cartilage, 2 - upper anterior nasal cartilage, 3 - lower anterior nasal cartilage, 4 - external opening of the nostril, 5 - oblique cartilage, 6 - anterior maxillary process, 7 - frontal fenestra, 8 - infraorbital fenestra, 9 - pterygoid process, 10 - transverse band of the cartilaginous bridge, 11 - middle band of the cartilaginous bridge, 12 - parietal window, 13 - main process of the quadrate cartilage, 14 - tympanic ring, 15 - interauditory bridge, 16 - oval window, 17 - articular process of the quadrate cartilage. Roman numerals indicate the exit points of the corresponding head nerves.

Rice. 5. The primary skull of an adult grass frog with the integumentary bones removed (a - top, b - bottom):
1 - oblique cartilage, 2 - girdle bone, 3 - infraorbital fenestra, 4 - frontal fenestra, 5 - parietal fenestra, 6 - pterygoid process (infraorbital arch), 7 - anterior auricular bone, 8 - lateral occipital bone.
From the material of the ear capsule in ontogenesis, the sound-conducting apparatus is separated - the lid of the ear (operculum auris) and the ear, or auditory, column (columella auris). The first is a small oval cartilaginous plate covering the posterior part of the oval window. The inner end of the ear column rests against the oval window, and the outer end against the eardrum. The middle part of the ear column ossifies (pars media = ossea). Its inner part (pars interna) is thickened, remains cartilaginous and is sometimes called “pseudoperculum”. The outer part (pars externa) of the ear column also remains cartilaginous, slightly widens downwards and is adjacent to the eardrum. From it goes upward a thin cartilage (processus ascendens), attached to the parotid cartilaginous crest (crista paraotica).
The cartilaginous base and roof of the primary skull, continuing forward into the orbital region, form a kind of tube. On its upper surface we see a large unpaired frontal window, or fontanelle (fenestra frontalis), behind which are paired parietal windows (fenestrae parietales). They are limited at the back by the already mentioned inter-auditory bridge, and in the middle they are separated by the middle band of the cartilaginous bridge (taenia tecti medialis).
The anterior end of each of the parietal fenestrae is limited by a transverse band of the cartilaginous bridge (taenia tecti transversalis). On the lateral surface of the posterior part of the orbital region, a large opening for the exit of the second nerve is clearly visible - the optic foramen (foramen opticum). Above and posterior to it there is a small opening for the IV nerve (foramen pro n. trochleari). The third cephalic nerve emerges between the posterior inferior angle of the optic foramen and the anterior auricular bone. More often it has one common opening (foramen oculomotorii) with the cerebral carotid artery, but sometimes instead there are two separate openings.
The posterior part of the nasal region is represented by the sphenoid-olfactory bone (os sphenethmoideum). It has the appearance of a belt and, located partly in the nasal region, covers the entire anterior part of the orbital region of the skull with a ring. The sphenoid-olfactory bone in an adult animal is an unpaired formation, whereas it is formed in the form of paired ossifications. At its posterior end there is a fossa, which makes up about a third of the anterior part of the braincase cavity. From this cavity, two olfactory canals (canales olfactorii) go forward through the body of the sphenoid-olfactory bone. Each of them unfolds forward like a funnel and serves for the passage of the first cerebral nerve. In front of the bone there is a belt of only cartilaginous mass, simply arranged in its posterior part and complicated in the front. The continuation of the bottom and roofs of the sphenoid-olfactory bone in front of it are the cartilaginous bases and the roof of the nasal region (solum = tectum nasale), and the bony septum separating the olfactory canals continues into the cartilaginous nasal septum (septum nasale). Thus, each olfactory canal continues forward, where it opens forward and sideways with the external opening of the nostril (apertura nasalis), and further forward with the main nasal window (fenestra nasobasalis). The external opening of the nostril is partially covered by the pterygoid (cartilago alaris) and oblique cartilages (cartilago obliqua). In the cartilaginous mass hanging over the main nasal window, there is a small frontal opening (foramen frontale). The paired lower anterior nasal cartilages (cartilagines praenasales inferiores) are attached below and closer to the midline of the body, and the upper anterior nasal cartilages (cartilagines praenasales superiores) lie higher and more laterally. The latter are essentially processes of oblique cartilage. Among the listed nasal cartilages, what is usually not noticed is the small horseshoe-shaped covering bone - the intranasal bone (os intranasale = sertomachillare). This bone lies below and to the side of the opening of the nostril.

Rice. 6. Primary skull of a young grass frog. Back view. Bone borders are omitted. The posterior half of the tympanic ring, the articular part of the quadrate cartilage and the cover of the ear were removed:
1 - foramen magnum, 2 - occipital condyle, 3 - jugular foramen, 4 - superior perilymphatic foramen, 5 - inferior perilymphatic foramen, 6 - oval window, 7 - 9 - inner, middle and outer part of the ear column, 10 - ascending process of the ear column, 11 - anterior part of the tympanic ring, 12 - part of the quadrate cartilage, 13 - pterygoid process, 14 - end of the main horn of the hyoid apparatus.
Moving on to large integumentary ossifications, we must first focus on the frontoparietal bones (ossa fronto-parietalia). On the skull of underyearlings, even in autumn, you can still see separate frontal (ossa frontalia) and parietal (ossa parietalia) bones. Subsequently, they merge on each side into a flat, rather wide bone, covering the posterior part of the sphenoid-olfactory bone in front, and part of the anterior auricular bone in the back. The former border between the frontal and parietal bones is located approximately at the level of the last quarter of the length of the orbit. In an adult animal, in its posterior part, each frontoparietal bone extends towards the posterior edge of the orbit. The lateral edges of the first two thirds of the frontoparietal bone are slightly curved downwards. The right and left bones are connected to each other along the midline by a sagittal suture (sutura sagittalis). In front of the sphenoid-olfactory bone is the paired nasal bone (os nasale). The degree of midline contact between the right and left nasal bones and the shape of its posterior margin are species specific. Even further forward is the paired premaxillary or intermaxillary bone (os praemaxillare = intermaxiflare). In it one can distinguish the facial (pars facialis), palatal (pars palatina) and dental (pars dentalis) parts. The facial part bears, closer to its inner end, an ascending process (processus ascendens), directed towards the nasal bone. In the middle of the palatine part of the premaxillary bone there is a more or less pronounced notch, due to which two processes are formed. The process (or rather the edge) lying outward is usually less developed. The jaw bone (osmacillare) forms the lateral contours of the skull and its anterior end is in contact with the premaxillary, and its posterior end is in contact with the quadratozygomatic. For a significant part of its length, this bone is underlain by the cartilaginous infraorbital arch. In the jawbone, the same three parts can be distinguished as in the premaxillary: facial (pars facialis), palatine (pars palatina) and dental (pars dentalis). The facial part of the jawbone is visible when viewing the skull from above. Approximately at the end of its first third there is a frontal process (processus frontalis ossis maxillaris), extended towards the nasal bone. The palatal part of the jawbone is a narrow horizontal strip visible when viewing the skull from below. The dental part serves as the boundary between the two already mentioned. The posterior continuation of the jawbone is the quadrato-jugal (os quadrato-jugaIe = quadrato-machillare) thin bone, which limits the posterior third of the oral fissure. As already mentioned, the bony head at the back of this bone is formed by ossification of a portion of the quadrate cartilage. Above the quadratojugal bone is the squamosal bone (os squamosum = tympanicum). In shape it resembles a hammer or the letter T and consists of a vertical body (corpus) and a horizontal part (pars horisontalis). The degree of development of the latter serves as a good species characteristic. Looking at the skull from below, we see an unpaired bone, somewhat reminiscent of a cross - this is the integumentary sphenoid or parasphenoid bone (os parasphenoideum = parabasale). Its body (corpus) underlies the cranium along the midline, while in the ear region this bone extends to the right and left along the lateral process (pars lateralis).
The pterygoid bone (os pterygoideum), which plays an important mechanical role in supporting both the cartilaginous and bony parts of the skull, is clearly visible from below, but also visible from above. The pterygoid bone consists of 3 parts. The pterygoid process (processus pterygoideus) goes forward to the jaw bone. The articular part (pars articularis) runs backward and outward, underlying the part of the quadrate cartilage below. Finally, a small main process (processus basalis) extends towards the integumentary sphenoid bone.
On the sides of the anterior end of the integumentary sphenoid bone lie the palatine bones (ossa palatina), each of which is a kind of bone stick located perpendicular to the longitudinal axis of the body. The upper surface of each palatine bone bears a kind of longitudinal groove.
Inside the space bounded behind by the palatine bones, and in front by the premaxillary bones and the anterior half of the maxillary bones, lies a paired vomer. The figure of this flat bone is complicated by several sharp processes. A number of teeth sit on the lower surface of the vomer. These teeth form a small row, the position of which is interesting for taxonomists. Often, according to tradition, the teeth sitting on the vomer are called “palatal teeth” (“dentes palatini”), meaning the palate as the entire upper roof of the oral cavity.

Rice. 7. Right jaw inside (a) and outside (b):
1 - articular surface of Meckel's cartilage, 2 - prearticular tubercle, 3 - cutaneous articular bone, 4 - coronoid process of cutaneous articular bone, 5 - dentary bone, 6 - Meckel-mental bone, 7 - anterior part of Meckel's cartilage.
The basis of the lower jaw of the frog is formed by Meckel's cartilage (cartilago Meckelii), which remains mostly unossified throughout life. The posterior part of Meckel's cartilage is expanded and forms an articular surface (facies articularis) for articulation with the quadrate cartilage. In front of the articular surface, Meckel's cartilage gives a small protrusion upward - the pre-articular tubercle (tuberculum praeglenoidale), and then forms a thinning cartilaginous rod. The anterior end of this rod (pars mentalis) usually ossifies into the Meckel-mental bone (os mento-meckelianum = mento-mandibulare). Overlying Meckel's cartilage are two integumentary ossifications: the cutaneous articular bone (os dermaticulare = goniaIe = operculo-angulare) and the dental (dentale). The first of them covers the posterior half of Meckel's cartilage from the inside and bottom. This bone gives rise upward to the coronoid process (processus coronoideus). The frog's dentary bone is devoid of teeth and, forming a kind of bone tube, covers most of the anterior half of Meckel's cartilage.
The jaw joint is formed by the articular surfaces of Meckel's and quadrate cartilages. The joint capsule is wide and extends forward to the prearticular tubercle of Meckel's cartilage, which provides not only wide opening of the mouth, but also a certain mobility back and forth.

Rice. 8. Sublingual apparatus:
1 - body, 2 - anterior process, 3 - manubrium, 4 - main horn, 5 - pterygoid process, 6 - posterolateral process, 7 - mid-posterior process.
The hyoid apparatus (cartilago hyoidea) develops from the gill skeleton of the tadpole and is difficult to understand without regard to ontogeny. It remains mostly cartilaginous throughout the animal's life. Its body (corpus hyoideum = corpus linguae) has the shape of a flat thin plate formed by the fusion of elements of the gill skeleton of the tadpole. Long main horns (cornua principalia = cornua hualia) extend from the anterior side of the body of the hyoid apparatus, each of which, curving backward and upward, is ultimately attached to the ear capsule. That segment of the main horns that is directed forward can be called the handle (manubrium). Approximately from the anterior end of the manubrium, an anterior process (processus anterior) extends, either running as in the figure, or curving, or even merging with the body. The anterior corners of the body of the hyoid apparatus are occupied by the pterygoid process (processus alaris), which develops from the cells of the horny hyoid cartilage of the tadpole. From each posterior corner of the body there extends a posterolateral process (processus posterolateralis), developing from the lower branchial cartilages. Finally, the mid-posterior processes (pr. postero-medialis = thyroidei) extend from the middle of the posterior edge, also developing from the lower branchial cartilages and ossifying for the most part.
Axial skeleton
In the process of phylogenesis of tailless amphibians, the vertebral column (columna vertebralis) was shortened, because while the body of a jumping animal is in the air (in “flight”), the long longitudinal axis of the body is mechanically disadvantageous. This shortening is achieved by successively moving forward the point of attachment of the pelvis: the last trunk vertebrae, in the process of phylogenesis, turn from sacral vertebrae into part of the caudal bone. It is possible that only the posterior part of the latter is homologous to the former caudal vertebrae. The disappearance of ribs also progresses in the phylogeny of the order: in round-tongued ribs there are still ribs in the adult state, in others they are formed and then during ontogenesis they merge with the transverse processes of the vertebrae, and in frogs their existence has not been proven at all.
Normally, the spine of a frog consists of 10 bones: 9 vertebrae and the caudal bone. An additional (tenth) trunk vertebra (atavistic variation) is often found; It is much less common to have one vertebra less than normal (progressive variation). The dense abdominal part of each vertebra - its body (corpus vertebrae) includes inside the remains of the notochord. Vertebrae from the second to the seventh have a body that is concave in front and convex at the back (anteroconcave type, vertebra procoela). These articular surfaces of the vertebra are covered with a layer of hyaline cartilage. Bone plates rise from the sides of the vertebral body (one on each side), which limit the nerve, or brain, canal (canalis vertebralis) and merge from above, forming the vertebral arch (arcus vertebrae = arcus neuralis). On the dorsal side, the vertebral arch bears a small unpaired spinous process (processus spinosus).
Most vertebrae also have a pair of anterior and a pair of posterior articular, or oblique, processes (pr. obliqui = zygaporhuses), as well as a pair of transverse processes (processus transversi), diverging more or less horizontally to the sides. In the anterior articular processes, their articular surfaces face upward and slightly to the sides, while in the posterior ones, they face downward and slightly towards the middle. The first vertebra deviates from the described typical case more than others. It is devoid of transverse processes (observed only as a rare atavistic anomaly) and the anterior pair of articular joints. The arch of the first vertebra usually does not completely ossify in the area of the spinous process, and the latter is in its infancy. The body of the first vertebra is extended forward into the unpaired interarticular process (tuberculum interglenoid). Previously, this process was called “tooth-like,” but its similarity to that of higher vertebrates is purely convergent. The eighth vertebra differs from typical ones only in that its body is biconcave (vertebra amphicoela). The ninth vertebra is the sacral vertebra (vertebra sacralis). Its body is convex in front and equipped with two convex tubercles at the back. The transverse processes of the sacral vertebra, articulating with the pelvis, are somewhat more expanded than the others, and are directed slightly upward and backward.
Rice. 9. Green frog spine from above. Roman numerals indicate the order of the vertebrae:
1 - interarticular tubercle, 2 - spinous process of the first vertebra, 3 - transverse process of the second vertebra, 4 - oblique process of the third vertebra, 5 - transverse process of the sacral (ninth) vertebra, 6 - tail, 7 - cartilaginous part of the tail.
Rice. 10. Green frog spine from below. Roman numerals indicate the order of the vertebrae.

Rice. 11. Fourth vertebra in front:
1 - vertebral body, 2 - medullary canal, 3 - spinous process, 4 - oblique process, 5 - transverse process.

Rice. 12. First vertebra in front:
1 - brain canal, 2 - neural arch.

Rice. 13. Sacral vertebra from behind.
Rice. 14. Tail in front:
1 - medullary canal, 2 - articular fossa, 3 - crest of the caudal bone.
The caudal bone, or tail (os coccygis=urostyI), is an unpaired long saber-shaped bone. If viewed from the front, one can see its body bearing two articular fossae (сavitas glenoidalis) for articulation with the convexities of the posterior surface of the body of the sacral vertebra. The body of the caudal bone continues backwards in the form of a thinning rod, which at the very end on dry skeletons has a tube-shaped depression. On fresh skeletons this depression is filled with cartilage, extending somewhat further back. Above the body, when viewed from the front, a short medullary canal is visible. The dorsal arch above it forms a spinous process, stretching back along the dorsal part of the caudal bone in the form of a ridge (spina ossis coccygei). The medullary canal opens on the lateral sides at the beginning of the ridge (Fig. 60) with a small opening - the caudal canal (canalis coccygeus). At the outer edge of the opening of the caudal canal there is (in green frogs more often than in brown frogs) a small triangular protrusion (processus rudimentalis), which is a rudiment of the transverse process. Cases of atavistic deformities have been described in which there were clearly defined transverse processes on the caudal bone.
Individual vertebrae articulate with each other through the anterior and posterior surfaces of their bodies and articular processes. These joints are strengthened thanks to the longitudinal abdominal (ligamentum longitudinale ventralе) and dorsal (lig. long, dorsale) ligaments, as well as thanks to the intercrucal (ligamenta intercrucalia) and interspinous (lig. interspinalia) ligaments. On the right and left, between each pair of upper arches, directly below the articular processes, there is an intervertebral foramen (foramen intervertebrale). The mobility of the spine is relatively limited. On the contrary, the movements of the caudal bone reach significant proportions. The back of a sitting frog seems humpbacked, but this false impression is created mainly due to the position of the tail bone and pelvis, while the spine itself remains completely straight at rest.
Rice. 15. Green frog tail on the right side:
1 - bone ridge, 2 - ventral edge, 3 - caudal canal, 4 - rudimentary process, 5 - cartilage.
Forelimb belt
The forelimb girdle consists of two symmetrical halves connected to each other on the ventral side along the midline of the body. In its entirety, this belt is a kind of semi-ring, the free ends of which are bent onto the dorsal side of the animal. On the border between the dorsolateral, otherwise scapular, and abdominal parts, the articular fossa of the humerus is located. The scapular part consists of the suprascapula and the scapula itself. The suprascapula occupies the outermost end of the semiring. This is an irregularly quadrangular plate, thinning towards its distal edge. Three sides of it remain free, and the fourth (the shortest) is attached to the scapula by means of a synchondrosis, a thin strip of hyaline cartilage. The anterior and proximal part of the suprascapula consists of bone, then there is a zone of calcified cartilage, and its dorsal, widest edge is covered with a thin strip of hyaline cartilage. Some believe that a small bony thickening on the anterior edge of the suprascapula is homologous to the cleithrum of stegocephalians. In some species

Rice. 16. Belt of the forelimbs of the green frog; straightened, ventral view:
1 - suprascapula, 2 - synchondrosis, 3 - scapula, 4 - acromial part of the scapula, 5 - acromion, 6 - articular fossa, 7 - periarticular cartilage, 8 - cartilaginous part of the presternum, 9 - bony part of the presternum, 10 - clavicle, 11 - corvid bone, 12 - supravranal cartilage, 13 - bony part of the sternum, 14 - cartilaginous part of the sternum.
(Rana natatrix) cleithrum is visible as a separate formation. The scapula has the shape of an elongated quadrangular bone plate. Its distal edge, associated with the suprascapula, is straight. Then the body of the scapula is somewhat narrowed, and further towards the midline of the body it expands again, forming two processes - the acromial (pars acromialis) and the articular (pars glenoidalis). The latter is clearly visible only from the inside of the belt, because it is directed inward. The proximal part of the scapula is connected to other parts of the pectoral girdle by cartilage. In general, the basis of the abdominal part of the girdle of the forelimbs is a cartilaginous “frame”. The anterior (head) crossbar of the frame is a thin anterior cartilage (cartilago procoracoidea), covered in front and on the ventral side by an overhead bone - the clavicle (clavicula). The rear crossbar of the frame ossifies in the form of a large corvid bone (os coracoid). On the side of the midline of the body, the frame is closed by the supravraneal cartilage (cart, epicoracoidea), and on the outside - by the periarticular cartilage (cart, paraglenoidalis). The angle of periarticular cartilage that projects forward between the distal end of the clavicle and the proximal end of the scapula is called the acromion. The supravranial cartilages of the right and left halves of the pectoral girdle are connected to each other in such a way that they allow some bending along the seam separating them, but they do not overlap each other, like in toads. The corvid bone is more or less constricted in its middle part; its proximal end (extremitas medialis) is wider than the distal one (extr. scarularis). The lower part of the latter takes a small part in the formation of the glenoid cavity of the humerus and therefore bears a small articular fossa (fovea acetabuli). The end of the clavicle (extremitas sternalis) directed towards the midline of the body is relatively straight, while the outer end (extr. scapularis) is curved and widened. In old frogs, progressive ossification of the listed cartilaginous formations is observed. In general, it is possible to clearly distinguish the boundaries of all the described formations only on the macerated skeletons of younger frogs.
An unpaired presternum (episternum = omosternum), consisting of a posterior bone (pars ossea) and anterior cartilaginous (pars cartilaginea) parts, runs forward from the junction of the supravraneal cartilages. The sternum (sternum) is also unpaired and also consists of bone (pars ossea) and cartilaginous (pars cartilaginea = “xiphisternum”) parts, but is directed not forward, but backward from the pectoral girdle.

Rice. 17. The lower part of the right half of the pectoral girdle of a green frog, visible from the inside:
1 - suprascapula (part), 2 - synchondrosis, 3 - scapula, 4 - acromial process of the scapula, 5 - articular part of the scapula, 6 - acromion, 7 - periarticular cartilage, 8 - clavicle, 9 - anterior wound
cartilage, 10 - corvid bone.
Forelimb
In an adult frog, the skeleton of the free limb has one bone in the shoulder, one in the forearm, and 21 in the hand.
The humerus (os humeri) articulates with the fossa of the shoulder girdle through the upper articular head (caput humeri), which has the shape of a ball. Part of this ball is cut out and forms the fovea capitis. Adjacent to the abdominal wall of the fossa is the ventral or deltoid crest (crista ventralis = deltoidea), which lies on the body of the humerus. Close to the upper articular head, located slightly below it and on the inner (medial) side, there is a small median tubercle (tuberculum mediale), covered, like the head, with calcified cartilage (Fig. 64). Down from the median tubercle, the process of the median tubercle (spinatuberculiinedialis) descends, approximately to the middle of the body of the humerus.
In the structure of the body of the humerus itself, there is a significant difference between the sexes: in mature males, a high median ridge (crista medialis) stretches along its median surface, descending to the very bottom of the bone and increasing at the time of spawning. In addition, males have a low lateral ridge (crista lateralis) on the outer surface of the lower part of the bone.
At the lower (distal) end of the humerus there is a hemispherical lower articular head (eminentia capitat a). On the ventral side in front of it there is a small abdominal fossa (fossa cubitalis ventralis). On one side of the lower articular head there is a median, or ulnar, process (epicondylus medialis = ulnaris), and on the other - a lateral, or radial (epicondylus lateralis = radialis). Below the median process, or olecranon, is the ulnar articular surface (condylus ulnaris). On the fresh skeleton, it can be seen that the articular capsule covers the proximal part of the humerus to the level of the median tubercle. The joint is strengthened by two scapulohumeral ligaments (the more powerful ligamentum scapulc-humerale mediale and the weaker lig. scapulo-humerale laterale) and one corvid-humeral ligament (lig. coraco-humerale). The joint allows movement along three main axes.

Rice. 18. Right humerus of a female green frog. Ventral view:
1 - superior articular head, 2 - fossa of the head, 3 - ventral crest, 4 - abdominal fossa, 5 - lateral process, 6 - inferior articular head, 7 - median process, 8 - ulnar articular surface.

Rice. 19. Right humerus of a male green frog. View from the inside:
1 - superior articular head, 2 - median tubercle, 3 - ventral ridge, 4 - process of the median tubercle, 5 - median ridge, 6 - abdominal fossa, 7 - inferior articular head, 8 - ulnar articular surface.
The forearm bone (os antibrachii) of the frog, judging by the shape of its distal end and the presence of a median groove (sulcus intermedius), is the result of the fusion of two bones - the radius and the ulna (ulna), fused in a semi-pronated position. At the proximal end of the forearm bone there is an articular fossa to receive the inferior articular head of the humerus. At the back, this fossa is bounded by the unpaired olecranon process, and in front by the unpaired coronoid process (processus coronoideus). At the distal end of the forearm bone, a smaller articular head formed by the ulna and a larger one formed by the end of the radius are clearly distinguishable. The movements of the elbow joint are limited by the aforementioned ulnar and coronoid processes, and a short, strong lateral ligament (ligamentum laterale) and a rather long median (lig. mediale) take part in strengthening it.
The interpretation of the skeleton of the frog's hand is complicated by theoretical disagreements. Old authors accepted that the first finger on the forelimb of the frog was lost. Newer studies have shown, however, that we must talk about the complete disappearance of the fifth finger. Accordingly, the skeleton of the nuptial callus of males should be considered not as a rudiment of the first finger, but as a rudiment of the finger that lay in front of the first (praeroIlex). The nomenclature of other bones will look different in this case. In this book, the second view is adopted as being in better agreement with both the structure of the limbs of tailed amphibians and paleontological data.
The six elements of the wrist are usually composed of somewhat calcified cartilage, but they can ossify with age. A verbal description of the shape of the carpal bones cannot give a clear idea of them. The description should be compared with drawings and preparations. Directly adjacent to the distal articular surfaces of the forearm bone are the ulnar carpal (ulnare = “ruramidale”) and the radial carpal (radiale = “lunaturm = “semilunar”). To the side and slightly in front of the latter is the central one (centrale-“navisulare”). The first authors tried to give these formations names (given in quotation marks) taken from human anatomy. Now no one doubts the impossibility of such homologization, but even to this day there remain some disagreements in understanding the morphological significance of individual elements. Thus, the intermediate carpal (intermedium) is not expressed in the frog, but exists in caudates - some authors believe that it merged with the ulnar carpal of the frog, and others - with the radial carpal. The question of the central one is no less complex: tailed animals have two or three central ones, but the frog has only one. It has been suggested that one of the central ones has merged with the radial carpal, but one can also speak of simply the disappearance of one of the central ones. The second (distal) row of the wrist is made up of three separate formations - various extreme carpal bones (carpalia distalia). At the base of the first finger there are two of them.

Rice. 20. Right forearm bone of the green frog:
1 - ulnar process, 2 - coronoid process, 3 - intermediate groove, 4 - ulna, 5 - radius.
Old authors considered them the first and second carpals, respectively, but now we have to change the count: we should talk about the carpal of the first (carpale praepollicis = “trapezium”) and the carpal of the first (carpaleI = “trapezoides”) fingers. The extreme carpal bones of the remaining (II-IV) fingers are fused together. Some believe that a second central one has grown to them. Between the ulna and radial carpal bones there is an intercarpal foramen (foramen intercarpale).
On the palmar surface of the ulnar carpal bone there is a longitudinal ridge (crista carpi longitudinalis), below which there is a small rear opening of the palm (foramen carpi posterius). on the back side there is a dorsal fossa of the hand (fovea dorsalis carpi), and on the palmar side there is a transverse ridge (crista trans versa carpi). The latter limits the distal part of the large palmar fossa (fovea volaris carpi), formed by the relief of a number of bones. Its lateral edge limits the already mentioned longitudinal crest of the ulnar carpal. Directly below the transverse ridge is the anterior opening of the palm (foramen carpi anterius).
Adjacent to the distal row of carpal bones are five metacarpals (metascarpalia). The extreme medial one is a rudiment of the metacarpal bone of the pre-first finger, and is noticeably more developed in males than in females. In females it is often just a small cartilage, while in males it is ossified to a certain extent. The carpal bones of the remaining fingers (metacarpalia I-IV) are simple cylindrical bones with epiphyses made of calcified cartilage.

Rice. 21. Skeleton of the right forearm and hand of a male green frog. View from the back (dorsal) side. Roman numerals indicate the order of the fingers:
1 - tuberosity of the first metacarpal bone, 2 - metacarpal bone of the first finger, 3 - carpal bone of the first finger, 4 - carpal bone of the first finger, 5 - dorsal fossa of the hand, 6 - carpal bone of the II-IV fingers, 7 - central bone, 8 - radial carpal bone, 9 - intercarpal foramen, 10 - ulnar carpal bone, 11 - radius bone,
The proximal part of the carpal bone of the first finger is expanded, and on its distal part there is a special tuberosity (tuberositas) in males.
Phalanges (phalanges digitorum) are present in fingers I-IV. Their number is 2, 2, 3 and 3, respectively. The longest finger is III. In our frogs, the terminal phalanges do not give noticeable expansion.
Among the tendons of the extensor carpi radialis there is cossal sesamoid cartilage.

Rice. 22. Skeleton of the right hand of a male green frog. View from the palm and somewhat from the inside. Roman numerals indicate the order of the fingers:
1 - tuberosity of the first metacarpal bone, 2 - metacarpal bone of the pre-first finger, 3 - carpal bone of the pre-first finger, 4 - carpal bone of the II-IV fingers, 5 - transverse ridge, 6 - anterior opening of the palm, 7 - central bone, 8 - radial carpal bone, 9 - intercarpal foramen, 10 - ulnar carpal bone, 11 - posterior foramen of the palm, 12 - longitudinal ridge, 13 - ulna, 14 - median groove, 15 - radius.

Rice. 23. The pelvis of a green frog. Bottom view:
1 - wing of the ilium, 2 - ilium, 3 - pubic cartilage, 4 - acetabulum, 5 - anterior pelvic spine.
Hind limb belt
If we consider the frog's pelvis from above or below, it looks like a Latin five - a vertical circle from which two branches extend forward.
This formation arose as a result of two processes: firstly, the convergence of the points of application of force of the hind limbs (important when jumping and swimming), and, secondly, as a result of the transfer of the attachment of the pelvis to the spine forward (shortening the long axis of the body is beneficial when moving in the air ). The morphological response of the body to the first circumstance was the convergence and fusion of the bones of the right and left halves of the pelvis into the mentioned circle, and to the second - the extension of the wings of the ilium forward.
The iliac bones (ossa ilei) are fused with their bodies, which take part in the formation of the acetabulum (acetabulum). Each bone has its own wing (aIaossisilei), the end of which remains cartilaginous and on which one can distinguish a blunt ventral (margo ventralis) and a sharp dorsal (margo dorsalis) edge (Fig. 69). The latter bears an upper process (processus superior) at its posterior end. The junction of the wing of the ilium with the body of the bone is called the wing root (radix alae).
The right and left ischiums (ossa ischii) are separate only in the tadpole. Their anterior edge takes part in the formation of the acetabulum.
In place of the pubic bone there is only cartilage, which is better called residual or pubic (cartilago remanens = “ocrubis”). The pubic cartilage holds the ilium and ischium together. The marginal cartilage (cartilago marginalis) extends from it, covering the entire sharp part of the ischium and continuing to the anterior (spina pelvis anterior) and posterior (spina pelvis posterior) spine. In very old individuals, calcification of the pubic cartilage is observed.
Species and sex differences in the structure of the pelvis have been established. The ridge running along the root of the ilium wing is sharper and more noticeable in the grass frog than in the green frog. Viewed in profile, the pelvic “discs” have different shapes: in the grass frog it is rounded and smaller. The anterior process of the ilium wing is more developed in the grass frog.
The articular prominence of the ischium is larger in the green frog. The pelvis of a male grass frog is shorter than that of a female: in a female, the total length of the pelvis divided by the length of the femur gives one or more. In a green frog, the pelvis can be recognized as that of a female if the total length of the pelvis divided by the length of the tibia gives 1.02 or more.
Hind limb
In an adult frog, the skeleton of the free hind limb has one bone in the femur, one in the tibia, and usually 25 formations in the tarsus.
Some of the latter remain cartilaginous for life.
The femur (os femoris) is one of the longest bones in the frog skeleton. Its middle part (diaphysis) is slightly S-shaped. The reasons for this shape become clear if we take into account the position that the femur occupies in the entire skeleton of the animal, since in this case the distal end of the bone is directed forward. At the proximal end of the bone there is a nearly spherical head of the femur (caput femoris), which on the whole skeleton enters the acetabulum of the pelvis. On the dorsal lateral surface near the head itself there is a small tubercle - trochanter for attachment of the external iliac muscle. The trochanter is separated from the head by the groove proper for the internal obturator muscle (sulcus pro m. obturator internus). On the mid-ventral side of the head there is a small fossa (fovea capitis).

Rice. 24. Pelvis of a green frog. Left view:
1 - wing of the ilium, 2 - its dorsal edge, 3 - its ventral edge, 4 - superior process, 5 - root of the wing, 6 - body of the ilium, 7 - anterior pelvic spine, 8 - posterior pelvic spine, 9 - acetabulum , 10 - pubic cartilage, 11 ischium, 12 - marginal cartilage.
The distal end of the femur bears an articular surface in the form of a transverse ridge. The shape of the latter allows us to distinguish between the external or lateral (condylus lateralis) and internal (condylus medialls) condyles.
The tibia bone (os cruris) is approximately equal in length or often longer than the femur and is then the longest bone of the frog skeleton. Longitudinal “intermediate” grooves on it (sulcus intermedius), especially clearly visible in the distal parts, show that this bone is the result of the fusion of two bones - the tibia and fibula. The nature of the frog's movement does not require rotation of these bones around each other, while their fusion increases strength. Transverse sections of the tibia bone show that there is a single bone marrow canal only in its middle part, and closer to the ends it is double. Both on the upper and lower surfaces of the tibia bone there is one hole in the middle part, which are connected to each other by a canal. On macerated bone, you can thread a bristle through them through the bone. The anterior tibial artery passes through this canal. This serves as further evidence of the fusion of bones that occurred in phylogenesis, for, as is known, for example, from human anatomy, the mentioned artery, after its departure from the popliteal, passes to the front side of the leg between two bones - the tibia and tibia. The shape of the proximal articular surface is quite complex. At the back it bears two condyles - the lateral (condylus lateralis) and the internal, or median (condylus medialis), separated by the intercondylar fossa (fossa intercondyloidea). Three grooves for the tendons of the muscles are visible in front: the peroneus, tibialis longus anterior and rectus tibia brevis. The distal end of the tibia bone bears on the dorsal side a rather deep groove for the tendon of the posterior tibialis muscle (sulcus pro m. tibialis posticus), and on the other side there is a flat capitate eminence (eminentia capitata).
Rice. 25. Right thigh bone of a green frog. Rear view and some outside view:
1 - head of the femur, 2 - trochanter, 3 - groove of the internal obturator muscle, 4 - internal condyle, 5 - external condyle.
Rice. 26. Green frog right shin bone. Ventral view:
1 - tibialis minor, 2 - tibia major, 3 - groove of the peroneal muscle, 4 - groove of the anterior tibialis longus muscle, 5 - groove of the short rectus tibia muscle, 6 - opening of the anterior tibial artery, 7 - intermediate groove, 8 - capitate eminence.
Rice. 27. Green frog right shin bone. View from above:
1 - intercondylar fossa, 2 - median condyle, 3 - lateral condyle, 4 - opening of the anterior tibial artery, 5 - intermediate groove, 6 - groove of the posterior tibial muscle.
The frog's tarsus (basiоdium) is characterized by a sharp difference between the elements of the distal and proximal rows. The former are largely underdeveloped, while among the latter two elements—the tarsal tibia (tibiale) and the tarsal tibia (fibulare)—are so highly developed that they imitate the bones of the lower leg. Some authors call the new limb segment formed due to the described circumstance “secondary tibia” (“crus secondarius”). When jumping, the frog's hind limb goes into the air segment by segment, starting from the thigh and ending with the outer phalanges of the fingers. The more segments there are, the more elastic the repulsion occurs and, apparently, the more fully the muscular force is used. This is likely an explanation for the formation of the “secondary tibia.” The tarsals, the tibia (tibiale = “talus” or “astragalus” in older authors) and the tibia (fibulare = “calcaneus”) are fused by their proximal and distal epiphyses, but in the middle part between them there is a large interosseous space (spatium interosseum). On the underside of the distal epiphysis of the tarsal tibia there is a trochlear process (processus trochlearis), hook-shaped. The intermediate bone (intermedium) in the frog is not expressed. Apparently, it has merged with the tarsal fibula. The distal row of tarsal elements is usually represented by calcified cartilage, the number of which varies from species to species. Thus, in the sharp-faced frog the first outermost one (tarsaleI) is not expressed, whereas in the grass and green frogs it is present. It is not surprising that a number of authors do not mention its existence at all. The extreme elements corresponding to the second and third fingers are fused together into a small bone (tarsale II + III). A continuation of the series of extreme elements is the additional ligament of the metatarsus (ligamentum tarsi supplens), which is sometimes considered as a remnant of the disappeared IV and V elements (old authors looked for a rudiment of the “os cuboideum” in it). The central one (centrale = “navisulare”) is well defined and is located between the end of the tarsal tibia and the base of the skeleton of the internal calcaneal tubercle.

Rice. 28. Skeleton of the right leg of a green frog from the dorsal side:
1 - tarsal fibula, 2 - tarsal tibia, 3 - interosseous space, 4 - extreme tarsus II+III, 5 - extreme tarsus I, 6 - central, 7 - skeleton of the internal calcaneal tubercle, 8 - additional ligament of the metatarsus. The metatarsals are indicated by Roman numerals.

Rice. 29. Skeleton of the right leg of a green frog from the sole side:
1 - sesamoid cartilage, 2 - tarsal tibia, 3 - tarsal tibia, 4 - interosseous space, 5 - trochlear process, 6 - extreme tarsus II+III, 7 - additional tarsal ligament, 8 - central, 9 - skeleton of the internal calcaneus mound. Roman numerals indicate the metacarpal bones.
The foot has 5 well-developed toes. Accordingly, the five metatarsal bones (ossa metatarsi) are well defined. The longest finger is the fourth and, accordingly, the number of phalanges (phalanges digitorum) will be (starting from the first): 2, 2, 3, 4, 3. All these are elongated cylindrical bones with articular surfaces at both ends. The terminal phalanges have articular surfaces only at the proximal end. In our species, the distal ends of the phalanges are slightly pointed or rounded. The skeleton of the internal calcaneal tuber appears to be homologous to the toe located in front of the first (praehalluh). The number of elements of which it is composed is quite variable: in the grass frog there are usually three cartilages in it, while in the sharp-faced frog there is often only one calcified element. With a significant degree of probability, the three mentioned elements can be considered as rudiments of a metatarsal bone and two phalanges of a reduced pre-first toe.
Below the proximal end of the tarsal tibia and below the articulation of the tarsal tibia and metatarsal fourth toe can be found by the sesamoid cartilage or bone (cartilago sesamoidea).
Literature used: P. V. Terentyev
Frog: Textbook / P.V. Terentyev;
ed. M. A. Vorontsova, A. I. Proyaeva. - M. 1950
Download abstract: You do not have access to download files from our server.
TOPIC 10. Amphibian SKELETON
SYSTEMATIC POSITION OF THE OBJECT
Subphylum Vertebrates, Vertebrata
Class Amphibians, Amphibia
Order Anurans, Anura (Ecaudata)
Representative - Lake frog, Rana ridibunda Pall.
MATERIAL AND EQUIPMENT
For one or two students you need:
1. A disassembled frog skeleton mounted on cardboard tablets.
2. Preparation needles - 2.
EXERCISE
Understand the structural features of the amphibian skeleton. Make the following drawings:
1. Frog skull from above.
2. Skull from below.
3. The spinal column and the pelvic girdle attached to it from above.
4. Belt of forelimbs (straightened) from below.
5. Skeleton of the forelimb.
6. Side view of the pelvic girdle.
7. Skeleton of the hind limb.
Additional task
Compare, without sketching, the skeletons of a tailless amphibian (frog) and a tailed amphibian (wet preparation).
DESCRIPTION OF THE SKELETON
The skeleton of amphibians, like other vertebrates, is divided into the axial skeleton (vertebral column), the skull (cerebral and visceral), paired limbs and their girdles.
In almost all parts of the skeleton, cartilage still plays a fairly important role.
Axial skeleton. The axial skeleton in amphibians is represented by the vertebral column (columna veriebralis; Fig. 54), consisting of ossified vertebrae; the notochord is usually reduced in adulthood. Compared to fish, the axial skeleton of amphibians consists of a larger number of sections.
Rice. 54. Axial skeleton and pelvic girdle of a frog (top view):
1 - cervical vertebra, 2 - trunk vertebrae, 3 - sacral vertebra. 4 - urostyle (fused caudal vertebrae), 5 - pelvic girdle, 6 - acetabulum
1. The cervical region (pars cervicalis; Fig. 54, 1) in all amphibians is represented by one cervical vertebra, which movably articulates with the skull using two articular platforms.
2. The trunk section (pars thoracalis; Fig. 54, 2) of the spine of frogs consists of 7 vertebrae (in tailed amphibians - from 14 to 63).
3. The sacral region (pars sacralis; Fig. 54, 3) in all amphibians is represented by one sacral vertebra, to the massive transverse processes of which the iliac bones of the pelvic girdle are attached (Fig. 54, 5).
4. The caudal region (pars caudalis) in the larvae of tailless amphibians consists of a fairly large number of individual vertebrae, which during metamorphosis merge into one tail bone - urostyle (urostyl; Fig. 54, 4). In tailed amphibians, 26-36 individual vertebrae are retained in the tail.

Rice. 55. Body vertebra of a frog
A - general view; B - longitudinal section:
1 - vertebral body, 2 - superior arch, 3 - canal for the spinal cord, 4 - spinous process, 5 - transverse process, 6 - articular process
The trunk vertebrae of most frogs are of the procoelous type: the vertebral body is concave in front, convex in the back (Fig. 55), however, the last trunk vertebra has an amphicoelous (biconcave) type of structure. Above the vertebral bodies are the upper arches (arcus neuralis; Fig. 55, 2), forming a canal for the spinal cord. On the dorsal side of the arch there is a small spinous process (processus spinosus; Fig. 55, 4). Paired transverse processes (processus transversus; Fig. 55, 5) extend from the superolateral surface of the vertebral body; In tailed amphibians, short ribs are attached to their ends; in tailless amphibians, there are no ribs. The vertebrae are connected to each other by the articulation of the vertebral bodies themselves (which is ensured by the procoelous type of their structure) and the connection of special paired articular processes (processus atricularis; Fig. 55, 6), located anteriorly and posteriorly at the base of the upper arch.
Compared to fish, amphibians are characterized by greater differentiation of the vertebral column into sections, a change in the shape of the vertebral bodies, and a stronger development of the articular processes. These transformations are associated with a terrestrial lifestyle and provide greater strength to the axial skeleton while maintaining its mobility, a strong connection with it of the pelvic girdle and allow some mobility of the skull in the vertical plane relative to the body (the ability to raise and lower the head).
Scull. The axial, or cerebral, skull of amphibians, like the skull of cartilaginous fish, is of the platybasal type: with a wide base and widely spaced orbits, between which the anterior end of the brain is located. The skull, compared to bony fish, retains a lot of cartilage, and the number of ossifications is relatively small.


Rice. 56. Frog Skull
A - from above; B - bottom; B - behind; D - lower jaw from above, the dotted line shows the cartilaginous areas of the skull
1 - lateral occipital bone, 2 - occipital condyle, 3 - anterior auricular bone, 4 - sphenoid-olfactory bone, 5 - nasal bone, 6 - frontoparietal bone, 7 - squamosal bone, 8 - parasphenoid, 9 - palatine bone, 10 - vomer, 11 - choana, 12 - palatoquadrate cartilage, 13 - premaxillary bone, 14 - maxillary bone, 15 - quadratozygomatic bone, 16 - pterygoid bone, 17 - Meckel's cartilage, 18 - geniomaxillary bone, 19 - dentary, 20 - angular bone, 21 - foramen magnum
In the cartilage of the occipital part of the skull, only paired lateral occipital bones (occipitale laterale; Fig. 56, 1) are formed, bordering the foramen magnum (foramen occipitale magnum; Fig. 56, 21). Each of them forms a condyle (condylus occipitalis; Fig. 56, 2) for articulation with the cervical vertebra. In the area of the auditory capsule, instead of five pairs of ear bones, characteristic of bony fish, amphibians have only one pair - the anterior ear bones (prooticum; Fig. 56, 3). In the anterior part of the brain skull, during ossification of the cartilage, an unpaired sphenoid-olfactory bone (sphenethmoideum, Fig. 56, 4) is formed, which has the appearance of a bone ring girdle.

Rice. 57. Cartilaginous tadpole skull:
1 - brain skull, 2 - palatoquadrate cartilage, 3 - Meckel's cartilage, 4 - branchial arches, 5 - jaw joint
The rest of the skull remains cartilaginous. It is strengthened by the integumentary (skin) bones. On top of the front part of the skull lie the paired nasal bones (nasale; Fig. 56, 5), which have an elongated triangular shape, then the paired frontoparietale bones (frontoparietale, Fig. 56, 6), fused from the frontal and parietal bones, and outward from the ear bones - scaly bones having a complex shape (squamosum; Fig. 56, 7). The bottom of the skull is covered by a powerful cruciform integumentary bone - the parasphenoid (parasphenoideum; Fig. 56, 8). In front of it also lie the paired integumentary palatine bones (palatinum; Fig. 56, 9) and paired vomers (vomer; Fig. 56, 10); Small teeth sit on the vomers. In front of the vomers there are paired internal nostrils - choanae (Fig. 56, 11).
The visceral region of the amphibian skull also retains a lot of cartilage. Throughout life, the palatoquadrate cartilage (cartilago palatoquadratum; Fig. 56, 12) is preserved, growing with its anterior end to the olfactory region of the cerebral skull, and with its posterior end to the base of the skull in front of the auditory capsule (Fig. 57, 2). Therefore, the skull of amphibians, like other terrestrial vertebrates, is autostylish according to the type of attachment of the jaw arch.
Adjacent to the palatoquadrate cartilage are the bones of the secondary upper jaw that arise in the skin: paired intermaxillare or praemaxillare; Fig. 56, 13), load-bearing teeth and maxillary bones (maxillare; Fig. 56, 14). Behind them, strengthening the posterior part of the palatoquadrate cartilage, the integumentary quadratojugal bone (quadratojugale; Fig. 56, 15) is formed on top, and the integumentary pterygoid bone (pterygoideum; Fig. 56, 16) is also formed below.
The primary lower jaw - Meckel's cartilage (cartilago Meckeli; Fig. 56, 17) also remains cartilaginous, only its very anterior end ossifies into small paired chin-maxillary bones (mento-mandibulare, Fig. 56, 18). They are joined by integumentary dental bones (dentale; Fig. 56, 19), which in amphibians lack teeth. The posterior part of the Meckel's cartilage is overgrown with a long integumentary angular bone (angulare; Fig. 56, 20) and several more small integumentary bones. Through the articular process of Meckel's cartilage, the lower jaw movably articulates with the posterior part of the palatoquadrate cartilage (Fig. 57, 5).

Rice. 58. Schematic section through the auditory region of the frog's head:
1 - brain, 2 - auditory capsule with semicircular canals, 3 - middle ear cavity, 4 - stapes, 5 - eardrum, 6 - eustachian tube, 7 - oral cavity
Complete reduction of the operculum in amphibians and the replacement of the hyostylistic type of attachment of the jaws with an autostylistic one lead to the loss of the main functions of the hyoid arch (strengthening the jaws, supporting the operculum).
Even in the ancestors of modern amphibians, the hyoid arch began to be reduced, and the cavity of the sputter (the remnant of the gill slits between the maxillary and hyoid arches) due to the transition to life in the air was transformed into the cavity of the middle ear (Fig. 58, 3). The upper element of the hyoid arch, the pendant (hyomandibulare), located next to the splatter, has turned into an auditory column-bone, or stapes (columellaa, or stapes; Fig. 58, 4). In modern tailless amphibians, the stapes has the appearance of a thin rod-shaped bone lying perpendicular to the brain skull under the squamosal and quadratojugal bones. One end of the stapes rests against the center of the eardrum (Fig. 58, 5), and the other against the oval window of the auditory capsule. This mechanism, which amplifies sound vibrations and provides the possibility of hearing in the air, is secondary reduced to varying degrees in some modern amphibians. Additional mechanisms that ensure the perception of sound waves propagating along a solid substrate are the lower jaw, as well as the transmission of sound vibrations along the blood trunks.

Rice. 59. Sublingual apparatus of the frog:
1 - body, 2 - horns
The lower element of the hyoid arch is the hyoid (hyoideum) and the gill arches that function in amphibian larvae transform into the hyoid apparatus during metamorphosis (Fig. 59). In tailless amphibians, it is a cartilaginous plate with two main layers of processes - horns. The anterior, longer horns (modified hyoids) are directed backward and upward and are attached to the walls of the auditory capsules of the brain skull. The hyoid apparatus strengthens the bottom of the oral strip: the muscles located between the branches of the lower jaw are attached to it.
It is assumed that the laryngeal cartilages are also transformed remains of the branchial arches.
Paired limbs and their belts. The limbs of amphibians, kak and the limbs of other classes of terrestrial vertebrates, represent in the diagram a system of levers movably connected to each other. The structural diagrams of the front and rear limbs are of the same type (Fig. 60):

Rice. 60. Scheme of the structure of paired limbs (left) of terrestrial vertebrates
A - forelimb; B - hind limb:
a - shoulder - thigh, b - forearm - lower leg, a - hand-foot;
1 - humerus, 2 - ulna, 3 - radius, 4 - wrist, 5 - metacarpus, 6 - phalanges, 7 - femur, 8 - tibia, 9 - fibula, 10 - tarsus, 11 - metatarsus, 12 - patella, I - V - fingers
Forelimb: Hindlimb:
I. Shoulder (humerus; Fig. 60, 1).
II. Forearm (antebrachium):
radius (radius; Fig. 60, 3),
ulna (ulna; Fig. 60, 2)
III. Brush (manus):
wrist (carpus; Fig. 60, 4),
metacarpus (metacarpus; Fig. 60, 5),
phalanges of the fingers (phalanges digitorum; Fig. 60, 6). I. Thigh (femur; Fig. 60, 7).
II. Tibia (crus): tibia (tibia; Fig. 60, 5),
fibula (fibula; Fig. 60, 9).
III. Foot (pes): tarsus (tarsus; Fig. 60, 10),
metatarsus (fig. 60, 11),
phalanges of fingers (phalanges digitorum; Fig. 60, 6)
The proximal part of the forelimb is the shoulder (humerus; Fig. 61, 7) - tubular bone; its middle part is called the diaphysis, and its thickened ends are called epiphyses. In amphibians, the epiphyses of the shoulder (and femur) remain cartilaginous.
The proximal end has a rounded head of the humerus (caput humeri; Fig. 61, 2), which enters the articular fossa of the girdle of the forelimbs; at the distal end there is a hemispherical surface for articulation with the bones of the forearm. The surface of the shoulder has ridges to which muscles are attached.
In tailless amphibians, the ulna (ulna; Fig. 61, 4) on the outside and the radius (radius; Fig. 61, 5) on the inside merge into a single forearm bone (antebrachium, Fig. 61, 3); the longitudinal groove shows the border of their fusion. In tailed amphibians these bones are independent.
The proximal ends of both bones form an articular fossa for connection with the shoulder; behind this fossa there is the olecranon process (Fig. 61, 6) of the ulna, which limits the extension of the limb.
The wrist (carpus, Fig. 61, 7) consists of two rows of small bones. Adjacent to the distal row of carpal bones are five elongated metacarpus bones (metacarpus; Fig. 61, 8). The phalanges of the fingers (phalanges digitorum; Fig. 61, 9) articulate with the distal ends of the metacarpal bones. In amphibians, the first (thumb) finger is greatly reduced and the hand ends with only four well-developed fingers.

Rice. 61. Forelimb and shoulder girdle of a frog:
1 - humerus, 2 - head of the humerus, 3 - forearm, 4 - ulna,
5 - radius, 6 - olecranon, 7 - wrist, 8 - metacarpus, 9 - phalanges, 10 - scapula, 11 - suprascapular cartilage, 12 - coracoid, 13 - articular cavity for the head of the humerus, 14 - procoracoid cartilage, 15 - clavicle, 16 - sternum, 17 - presternum, I - reduced first finger, II - V - well-developed fingers
The girdle of the forelimbs, or shoulder girdle, in amphibians, as well as in shark fish, lies in the thickness of the muscles of the body, connecting it with the axial skeleton. The scapula (scapula; Fig. 61, 10) is formed from the upper (dorsal) scapular part of the primary girdle; its uppermost part remains cartilaginous in the form of a wide suprascapular cartilage (cartilago suprascapularis, Fig. 61, 11). On the anterior outer surface of the suprascapular cartilage in some tailless amphibians there is a small ossification - a remnant of the cleithrum of fish-like ancestors. The ossified coracoid part of the girdle turned into a powerful coracoid bone (coracoideum; Fig. 61,12), together with the scapula, forming an articular cavity for the head of the humerus (Fig. 61, 13). Anterior to the coracoid, behind a small opening, lies a cartilaginous procoracoid (cartilago procoracoidea; Fig. 61, 14), on which lies a thin covering bone - the clavicle (clavicula; Fig. 61, 15). The unossified cartilaginous inner ends of the coracoids and procoracoids of the right and left sides fuse together along the midline. Behind the coracoids there is a bony sternum (sternum, Fig. 62, 16) with a cartilaginous posterior end. Anterior to the procoracoids protrudes the presternum (praesternum; Fig. 61, 17), also with a cartilaginous end. In the girdle of the forelimbs of caudate amphibians there is noticeably more cartilage, and the ossifications are smaller; The collarbones often do not develop.
The shoulder girdle serves as a support for the forelimbs and the attachment point for the muscles that control them.
The rib cage in amphibians does not develop: the sternum does not articulate with the ribs.

Rice. 62. Hind limb (A) and pelvic girdle (B) of a frog from the side:
1 - femur, 2 - femoral head, 3 - tibia, 4 - tibia, 5 - fibula, 6 - tarsus, 7 - tibiale, 8 - fibulare, 9 - metatarsus, 10 - phalanges, 11 - rudiment VI finger, 12 - ilium, 13 - ischium, 14 - pubic cartilage, 15 - acetabulum, I - V - fingers
The hind limb has an elongated tubular bone - the femur (femur; Fig. 62, 1), the proximal part of which ends with the head (Fig. 62, 2), which enters the acetabulum (Fig. 62, 15) of the pelvic girdle. The tibia (tibia; Fig. 62, 4) and fibula (fibula, Fig. 62, 5) bones of tailless amphibians merge into a single tibia bone (crus, Fig. 62, 3); in tailed amphibians they remain separated.
The proximal row of tarsal bones (tarsus, Fig. 62, 6) of tailless amphibians consists of two elongated bones that form an additional lever of the limb. The inner one is called tibiale (astragalus; Fig. 62, 7; adjacent to the tibial edge of the tibia), the outer one is called fibulare (calcaneus, Fig. 62, 8). The ankle joint is formed between the lower leg and these bones. From the distal row of tarsal bones in amphibians, only 2-3 small bones are preserved. The metatarsus (metatarsus; Fig. 62, 9) is formed by five long bones, to which the phalanges of the fingers (phalanges digitorum; Fig. 62, 10) are attached. The longest finger in frogs is IV. On the side of the I (inner) finger there is a small rudiment of the VI (“pre-first”) finger (praehallus; Fig. 62, 11).
The hind limb girdle, or pelvic girdle, in amphibians, as in all terrestrial vertebrates, consists of three paired elements; and all of them together form the articular acetabulum (acetabulum; Fig. 62, 15) for connection with the femoral head. The long, forward-directed iliac bones (ilium; Fig. 62, 12) are articulated at their ends to the transverse processes of the sacral vertebra (see Fig. 54). The lower part of the pelvic girdle in amphibians does not ossify and is represented by pubic cartilage (cartilago pubis, Fig. 62, 14). Behind it lie the paired ischiums (ischium; Fig. 62, 13).
In tailed amphibians, compared to tailless amphibians, there is much more cartilage in the pelvic girdle, and the formed bones are small.
CONCLUSION
Amphibians (Amphibia) are the first class of terrestrial vertebrates. However, representatives of the class still maintain a constant connection with water. This duality is clearly manifested in the characteristics of embryonic and postembryonic development. Eggs (spawn) can only develop in water (or in rare cases in an extremely humid environment). A larva emerges from the egg - a tadpole, which has clearly expressed signs of a typical aquatic animal: it has gills and gill arches supporting them, a two-chambered heart, one circle of blood circulation, paired limbs of the terrestrial type are absent, the main organ of movement is a powerful caudal fin, lateral line organs are developed etc. During metamorphosis (transformation), the larva loses some of the characteristics characteristic of aquatic animals and acquires features that ensure the transition to a terrestrial (or rather, terrestrial-aquatic) way of life: paired limbs of the terrestrial type appear, lungs develop, gills are reduced and the skeletal apparatus supporting them, the circulatory system is rebuilt - two incompletely separated circles of blood circulation are formed, etc.
The duality of organization as an adaptation to a terrestrial-aquatic lifestyle is also well expressed in adult individuals.
The terrestrial lifestyle is ensured by a number of structural features: greater differentiation of the spinal column into sections and a stronger connection of the vertebral bodies with each other (replacement of amphicoelous vertebrae with procoelous or opisthocoelous); the appearance of paired terrestrial limbs; increased complexity of the structure and greater strength of the limb girdles (in this case, a sufficiently strong connection of the pelvic girdle with the axial skeleton is already established); strong reduction of metameric muscles and its replacement by a fairly powerful complex complex of muscles; the appearance of eyelids (protecting the eyes from mechanical damage, preventing the cornea from drying out, etc.); formation of the middle ear cavity with the eardrum and the auditory ossicle - the stapes (ensuring the possibility of hearing in the air). A major role was played by the disappearance of the gills and the development of the lungs, larynx and choanae, which create the possibility of air breathing; the emergence of two circles of blood circulation; greater differentiation of the digestive system (high energy costs when maintaining the body in the air), etc.
The general lengthening of the hind limbs, the isolation of an additional lever in them (due to the sharp lengthening of the two proximal tarsal bones) and the possibility of a strong bend in the middle of the body at the junction of the branches of the iliac bones with the transverse processes of the sacral vertebrae - adaptations for jumping movement in tailless amphibians. Crawling tailed amphibians do not have these features. The fusion of two bones of the forearm and two bones of the lower leg into a single whole is associated with a sharp decrease in the need for rotational movements of the foot and hand when moving by jumping. In tailed amphibians, both the forearm and the lower leg consist of two independent elements, providing the rotational movements of the hand and foot necessary for crawling.
“Aquatic” structural features are manifested in a number of features: the relatively weak development of skeletal ossifications, the abundance of mucous glands in the skin (the mucus covering the skin reduces friction when moving in water, prevents the penetration of bacteria and fungi into the skin, etc.), preservation of the tail, often flattened laterally and bordered by a leathery fold (newts and other tailed amphibians), great similarity of the genitourinary system with most groups of fish, external fertilization characteristic of the vast majority of amphibian species, etc.
With a relatively small surface area of the lungs of amphibians, fairly powerful additional respiratory organs are required. Such an organ becomes the skin that is always moist (due to the abundance of mucous glands), easily permeable to moisture and gases, and partly the mucous membrane of the oral cavity. In an active pond frog, the lungs absorb up to 50% of the oxygen consumed by the body and release only about 14% of carbon dioxide; through skin respiration, up to 50% of oxygen is absorbed and up to 86% of carbon dioxide is released. In the grass frog, which leads a more terrestrial life, pulmonary respiration takes in up to 67% of oxygen and releases up to 26% of carbon dioxide, and through skin respiration, 33% of oxygen is absorbed and 74% of carbon dioxide is released. With an increase in the level of metabolism (increased general activity and all metabolic processes with increasing environmental temperature), the specific role of the lungs in providing the body with oxygen increases markedly. A decrease in environmental temperature causes a decrease in the level of metabolism. At the same time, skin respiration almost completely ensures both the saturation of the body with oxygen and the release of carbon dioxide, and the relative importance of the lungs in breathing sharply decreases.
This duality in the nature of breathing is explained not only by the insufficient development of the surface of the lungs and the imperfection of pulmonary ventilation (“swallowing” of air in the absence of a chest); it is necessary for the amphibian lifestyle of representatives of this class. It is this duality of the respiratory organs that provides amphibians with the opportunity to stay in water for a long time (up to wintering at the bottom of the reservoir of many species of anurans, when, with a sharp decrease in the level of metabolism, skin respiration completely meets all the body’s needs for oxygen and the release of carbon dioxide).
Using skin for breathing is possible only when it is easily permeable to moisture and gases. But such skin cannot protect the body from large losses of water (drying). Therefore, almost all types of amphibians inhabit only damp, damp areas, where the body loses less moisture and can always replenish its loss. Relatively few toads associated with water (overwinter on land and go into water bodies only to spawn) have thickened skin; this reduces the possibility of cutaneous respiration, which is compensated by an increase in the inner surface of the lungs. However, even in them, despite the thickening of the skin, the body loses up to 15-30% of water during the night hunting period. Some reduction in moisture loss (while maintaining skin permeability) in amphibians is helped by the mucus covering the skin.
Extensive subcutaneous lymphatic cavities serve as reservoirs of reserve water. Moisture loss is also reduced due to the reabsorption of water in the bladder, hind intestine and cloaca. Moisture loss decreases very sharply due to adaptive behavioral features: amphibians show increased activity only during hours of maximum air humidity (in clear weather - at dusk, as well as at night); they go to rest in burrows, where high humidity is maintained due to soil moisture.
The duality of the respiratory organs makes it impossible to completely separate the systemic and pulmonary circulations. But specific features in the structure of the heart and the blood trunks extending from it provide some separation of the blood flow, despite the fact that there is only one ventricle in the heart of amphibians, and there is an admixture of arterial blood in the superior vena cava. The development of muscular outgrowths of the walls of the ventricle reduces the mixing of blood, and the departure of the arterial cone from the right (more venous) side of the ventricle and the details of its internal structure (the sequence of origin of the arterial arches, the device of the spiral valve) allow more venous blood to be directed into the skin and lungs, more arterial - to the brain and sensory organs.
Greater differentiation of the digestive tract, compared to fish, leads to a slight increase in the intensity of food use. However, the rate of digestion in amphibians is low and depends on the ambient temperature. Food connections are quite simple; the range of feed used is small (only animals of relatively small size).
Amphibians, like fish, are characterized by variability in body temperature (poikilothermia): in amphibians it is usually only 0.5-1°C higher than the ambient temperature. Only during the period of highest activity (pursuing prey, avoiding danger) can body temperature exceed the ambient temperature by 5-7°C.
Poikilothermy causes a pronounced seasonal change in activity in amphibians of temperate and northern latitudes: when the air temperature drops to +5 - +8°C, all amphibians go into shelter (some species of frogs - into holes at the bottom of reservoirs; most species of tailless and tailed amphibians hide in rodent burrows, rotten tree roots, heaps of moss, etc.) and fall into a state of stupor. The geographical distribution of amphibians is also connected with this: most species of these animals are characteristic of the tropical zone. In the tropics, under relatively stable temperature conditions throughout the year, a state of torpor in a number of amphibian species is caused by a sharp decrease in air humidity (“hibernation” during the dry period of the year).
The very high dependence of amphibians on environmental humidity and temperature is reflected in the fact that weather conditions (in our latitudes - severe droughts in summer, severe frosts without snow in winter) often serve as the main cause of mortality and determine sharp fluctuations in the number of amphibians from year to year.
additional literature
Bannikov A. G., Denisova M. N. Essays on the biology of amphibians. M., 1956
Vorontsova M. A., Liozner L. D., Markelova I. V., Pukhelskaya E. Ch. Triton and the axolotl. M., 1952.
Gurtovoy N. N., Matveev B. S., Dzerzhinsky F. Ya. Practical zootomy of vertebrates. Amphibians, reptiles. M., 1978.
Terentyev P.V. Frog. M., 1950.
Terentyev P.V. Herpetology. M., 1961.
Shmalgauzen I.I. Fundamentals of comparative anatomy. M., 1947.
Shmalgauzen I. I. Origin of terrestrial vertebrates. M., 1964.
In the upper part of the body cavity lies a three-chambered heart (Fig. 38). The dark-colored atria and lighter ventricle are clearly visible.
On the sides of the heart lie dark gray thin-walled lungs. As a rule, they collapse when opened and are therefore difficult to see. It is necessary to note the thin-walledness of the lung sacs, the weak cellularity of their surface and the network of blood vessels in their walls.
Below the heart is a large three-lobed liver. A greenish-brown gall bladder is visible between the lobes of the liver.
Figure 38 – Dissected frog
1 – heart; 2 – lung; 3 – liver; 4 – gallbladder; 5 – stomach; 6 – pancreas; 7 – duodenum; 8 – small intestine; 9 – rectum; 10 – spleen; 11 – cloaca; 12 – bladder; 13 – kidney; 14 – ureter; 15 – right ovary (left ovary removed); 16 – fat body; 17 – right oviduct; 18 – uterine section of the oviduct;
19 – dorsal aorta; 20 – posterior vena cava; 21 – carotid artery;
22 – left aortic arch; 23 – pulmonary artery.
Under the liver, on the left side of the body, is the stomach, which passes into the duodenum. In the loop between the duodenum and the stomach, a small orange-yellow pancreas is attached to the mesentery.
The duodenum passes into the small intestine, which is curled into a ball. The rectum is very clearly defined. On the mesentery, approximately at the level of the anterior edge of the rectum, lies the burgundy-colored spleen. Above the rectum, at the point where it exits into the cloaca, there is a transparent, two-lobed bladder (often upon opening it is damaged, collapses and is difficult to see).
The kidneys are located on the dorsal side of the abdominal cavity and are covered by the intestines, and in female frogs, by the genitals.
Lifting the intestines (and ovaries in females) with tweezers, we will see the kidneys and the fat bodies (genital appendages) lying in front of them, which are represented by multi-lobed flat formations.
If the male is opened, then under the intestines we find a pair of oval testes. In a sexually mature female, the entire posterior part of the body cavity is occupied by ovaries filled with eggs (spawn) and long oviducts rolled into a complex ball. It should be emphasized that the reproductive system of females is usually so developed that it even covers the intestines, so to examine the latter it is necessary to move the ovaries and oviducts to the sides.
Compared to bony fish, the digestive system of amphibians is characterized by further complexity and differentiation.
The digestive tube begins with an oral slit leading into the oropharyngeal cavity (the latter was studied during an external examination of the frog). It should only be recalled that the tongue is placed in this cavity. The ducts of the salivary glands, which first appear in amphibians, open into it. However, in frogs these glands serve only to wet the food bolus and are not yet involved in the chemical processing of food.
The oropharyngeal cavity passes into a short but wide esophagus (Fig. 39), and the latter into a relatively voluminous stomach, which has a slightly curved shape.
The pyloric part of the stomach, bending strongly, passes into the duodenum, which is the beginning of the small intestine. As already indicated, the pancreas lies in the loop between the stomach and duodenum. The small intestine forms many bends and loops and smoothly passes into the large intestine, which ends in a clearly visible rectum. The rectum opens into the cloaca. The entire intestine is suspended from the walls of the cavity on special folds of the peritoneum - the mesentery.
The digestive glands - liver with gall bladder and pancreas - are well developed. The liver ducts, together with the gallbladder duct, open into the duodenum. The pancreatic ducts flow into the gallbladder duct, so this gland does not have independent communication with the intestines.

Figure 39 – Frog intestine
1 – esophagus; 2 – stomach; 3 – duodenum; 4 – small intestine; 5 – rectum; 6 – cloaca; 7 – place where the rectum flows into the cloaca; 8 – bladder.
The respiratory organs of amphibians are of a completely different type than those of fish. They are represented by light ones - two thin-walled oval-shaped bags with narrow lower ends (Fig. 38). The inner surface of the lungs is slightly honeycombed. However, as a result of the imperfection of the lungs (small oxidation surface), the skin plays an important role in breathing. For example, in green frogs, over 50% of the oxygen necessary for blood oxidation passes through the skin. In connection with pulmonary breathing, internal nostrils, or choanae, appear, connecting the nasal cavity with the oropharyngeal cavity.
The airways are very short due to the absence of the cervical spine. They are represented by the nasal and oropharyngeal cavities, as well as the larynx. The larynx opens directly into the lungs with two openings.
The circulatory system of amphibians has undergone significant transformations and is significantly different from that of fish. In connection with the appearance of the lungs, a second circle of blood circulation and a three-chambered heart arose (Fig. 40).
The frog has a three-chambered heart; it consists of the right and left atria and a ventricle. The right atrium is more voluminous - blood from the whole body collects through the veins into it, while the left atrium receives blood only from the lungs.
The ventricle is thick-walled, its inner surface is covered with numerous projections, between which there are pocket-like depressions.

Figure 40 – Diagram of an opened frog heart
1 – right atrium; 2 – left atrium; 3 – ventricle; 4 – valves that close the common opening leading from both atria to the ventricle; 5 – arterial cone; 6 – common arterial trunk;
7 – pulmonary cutaneous artery; 8 – aortic arch; 9 – common carotid artery; 10 – carotid gland; 11 – spiral valve of the arterial cone.
In addition to the indicated main parts of the heart, there is a venous sinus (sinus), which communicates with the right atrium, and an arterial cone extending from the right side of the ventricle.
Three pairs of arterial vessels (arterial arches) depart from the conus arteriosus. All three vessels (arches) of the left and right sides first go through a common arterial trunk, surrounded by a common membrane, and then branch (Fig. 40 and 41).
The vessels of the first pair (counting from the head) are called the carotid arteries. The carotid arteries carry blood to the head. These vessels depart from the common arterial trunk in the form of the common carotid arteries, each of which almost immediately splits into the external and internal carotid arteries (Fig. 41). At the site of their separation lies the carotid gland, which regulates blood pressure in the carotid arteries.

Figure 41 – Diagram of the arterial system of a frog
1 – ventricle; 2 – right atrium; 3 – left atrium;
4 – arterial cone; 5 – common carotid artery; 6 – aortic arch;
7 – subclavian artery; 8 – dorsal aorta; 9 – iliac artery; 10 – femoral artery; 11 – sciatic artery; 12 – enteromesenteric artery; 13 – pulmonary artery; 14 – cutaneous arteries;
15 – carotid gland; 16 – external carotid artery;
17 – internal carotid artery (arteries with venous blood are painted black, arteries with arterial and mixed blood are shaded).
Through the vessels of the second pair - the aortic arches - blood is directed to the back of the body. The arches go around the heart on the right and left sides, respectively, and merge under the spine into a common trunk - the dorsal aorta. Subclavian arteries depart from the aortic arches, carrying blood to the forelimbs (Fig. 41).
Through the vessels of the third pair - the pulmonary arteries - blood is sent to the lungs. A large cutaneous artery branches off from each pulmonary artery, through which blood is directed to the skin for oxidation (Fig. 41).
From the dorsal aorta, blood is carried through a series of arteries to the internal organs and hind limbs.
Venous blood (Fig. 42) from the anterior end of the body is collected through two pairs of jugular veins. The latter, merging with the cutaneous veins, which have already absorbed the subclavian veins, form two anterior vena cava. These veins carry mixed blood into the venous sinus, since oxygenated arterial blood moves from the skin through the cutaneous veins.
Blood from the hind limbs and back of the body moves through the iliac veins to the kidneys, where it passes through the portal system. The vessels leaving the kidneys merge to form the powerful posterior vena cava. Through the posterior vena cava, blood is directed to the venous sinus, from which it then enters the right atrium (Fig. 42).
From the intestines, blood is collected by the subintestinal vein, which flows into the liver, where the portal system functions. Blood also passes through the portal system of the liver from the abdominal vein, which carries it from the hind limbs. From the liver, blood flows through the hepatic veins into the posterior vena cava (Fig. 42).
From the lungs, blood moves through the pulmonary veins to the left atrium.
The blood circulation in the heart of a frog can be schematically represented as follows. Mixed blood enters the right atrium, and arterial blood (from the lungs) enters the left atrium. When the atria contract, blood flows through the common opening into the ventricle. This is where further mixing of the blood occurs. However, venous blood predominates in the right part of the ventricle, and arterial blood predominates in the left. The opening leading from the ventricle to the conus arteriosus is located on the right side of the ventricle. Therefore, when the ventricle contracts, the first portion of blood, containing more venous blood, enters the opening of the nearest pulmonary arch, the next portion, with a predominance of arterial blood, enters the aortic arch, and the portion with the least content of venous blood enters the carotid arteries.

Figure 42 – Diagram of the venous system of a frog
1 – venous sinus; 2 – external jugular vein; 3 – internal jugular vein; 4 – great cutaneous vein; 5 – subclavian vein;
6 – anterior vena cava; 7 – posterior vena cava; 8 – femoral vein;
9 – sciatic vein; 10 – iliac vein; 11 – renal portal system; 12 – subintestinal vein; 13 – portal system of the liver;
14 – hepatic veins; 15 – abdominal vein; 16 – pulmonary vein (veins with arterial blood are shaded).
The excretory organs (Fig. 43, 44) are represented in amphibians, as well as in fish, by the trunk kidneys (mesonephros). They are elongated, compact, reddish-brown bodies that lie on the sides of the spine. From each kidney a thin Wolffian canal stretches to the cloaca. In female Wolffrogs, the canal serves only as an excretory duct, or ureter, while in males it simultaneously serves as the genital duct, or vas deferens. In the cloaca, the Wolffian canals open with independent openings. It also opens separately into the cloaca and bladder. Urine enters first into the cloaca, and from it into the bladder. After filling the latter, through the same hole, urine is discharged again into the cloaca, and then out.
The reproductive organs of amphibians are represented by paired gonads. In males, these are oval-shaped testes, attached by a special mesentery to the anterior part of the kidneys (Fig. 43). Thin seminiferous tubules stretch from the testes to the kidneys. Sexual products from the testis are sent through these tubules to the bodies of the kidneys, then into the already known Wolffian canals and through them to the cloaca. Before flowing into the cloaca, the Wolffian canals form small expansions - seminal vesicles, which serve to temporarily reserve sperm.
The ovaries of females (Fig. 44) are thin-walled sacs, filled with eggs in adults. In the lateral parts of the body cavity there are highly convoluted light oviducts, or Müllerian canals. These genital canals are not directly connected to the ovaries; they open through small funnels near the lungs into the body cavity. Before flowing into the cloaca, each oviduct expands into the so-called “uterus”. Mature eggs fall out through breaks in the walls of the ovary into the body cavity, then are captured by the funnels of the oviducts and move along them to the cloaca. Thus, in females the excretory and reproductive ducts are completely separated.
The central nervous system in amphibians is represented, like in all vertebrates, by the brain and spinal cord (Fig. 45).

Figure 45 – Frog brain top and bottom
1 – cerebral hemispheres of the forebrain; 2 – olfactory lobe;
3 – olfactory nerve; 4 – diencephalon; 5 – visual chiasm; 6 – funnel; 7 – pituitary gland; 8 – optic lobes of the midbrain;
9 – cerebellum; 10 – medulla oblongata; 11 – spinal cord.
Compared to fish, the amphibian brain has a number of progressive features. This mainly concerns the forebrain, which is relatively larger than that of fish, its hemispheres are completely separated, and the nervous substance lines, in addition to the bottom of the lateral ventricles, also the sides and the roof, i.e. Amphibians have a true primary medullary vault, the archipallium.
The frog's brain consists of five sections (Fig. 45). In front is the forebrain, consisting of two elongated hemispheres separated by a deep fissure. In front of the hemispheres there is a common olfactory lobe, from which two olfactory nerves originate. Behind the forebrain is the diencephalon. On its roof is the epiphysis (endocrine gland). The midbrain is presented in the form of two rounded optic lobes. Behind the optic lobes lies the underdeveloped cerebellum. Immediately behind it is the medulla oblongata with the rhomboid fossa (the fourth cerebral ventricle). The medulla oblongata gradually transforms into the spinal cord.
On the lower side of the brain we find the optic chiasm, or chiasm, extending from the bottom of the diencephalon, the infundibulum and the pituitary gland.
Amphibian skeleton
Theoretical information:
11.1 General characteristics
11.2 Skeletal structure
general characteristics
Axial skeleton consists of vertebrae and is divided into 4 sections: cervical (1 vertebra), trunk (7 vertebrae), sacral (1 vertebra), caudal (vertebrae in tailless animals merge into the urostyle). In caudates, the total number of vertebrae reaches 200–300. There are 3 types of vertebrae in amphibians:
a) amphicoelous (in primitive legless amphibians);
b) opisthocoelous, the vertebral bodies are convex in front, concave in back (in tailed amphibians);
c) procoelous, the vertebral bodies are concave in front, convex in back (in tailless amphibians).
The superior arches are developed above the vertebral bodies, forming the spinal canal. At the base of the upper arches lie articular processes that articulate with similar processes of neighboring vertebrae; this increases the strength of the spinal column without reducing its flexibility. Below the articular processes there are transverse processes, unequally developed in different parts of the spine: in the cervical vertebra they are very poorly developed; in the trunk region, short ribs are attached to the developed transverse processes that do not reach the sternum (there is no rib cage); at the sacral vertebra the transverse processes are most developed; the iliac bones of the pelvic girdle are attached to them. In caudates, the lower arches of the caudal vertebrae form the hemal canal.
Scull retains a lot of cartilage. Amphibians are characterized by autostyly (the palatoquadrate cartilage of the upper jaw is fixedly attached to the cranium) and platybasality (the base of the cranium is wide, the brain is located between the orbits).
a) Brain skull:
The occipital region is represented by 2 lateral occipital bones bordering the foramen magnum. Each lateral occipital bone has a condyle for connection to the cervical vertebra.
The auditory section is represented by one pair of anterior auricular bones and integumentary squamosal bones.
The orbital region in caudates is formed by the oculosphenoid bone, and in tailless animals - by the sphenoid-olfactory bone.
The roof of the brain skull is formed by 2 pairs of bones: nasal and frontoparietal (in caudates, the frontal and parietal bones are separated).
The bottom of the brain skull is formed by a large cruciform parasphenoid, anterior to which are paired integumentary palatine bones and vomers on which small teeth sit.
The rest of the brain skull remains cartilaginous.
b) Visceral skull:
Jaw arch:
Upper jaw:
The palatoquadrate cartilage persists throughout life. A number of integumentary bones are adjacent to it: the premaxillary (intermaxillary) and maxillary in front and on the sides, the quadratozygomatic and squamosal in the back and above, the pterygoid in the back and below.
Lower jaw:
It is represented by Meckel's cartilage, covered with integumentary bones: the chin-maxillary bones in front, the main part covered by the dentary bones, and the long angular bones in the back. The jaw joint is formed by the articular process of Meckel's cartilage and the posterior portion of the palatoquadrate cartilage.
Hyoid arch:
The upper element of the hyoid arch - the hyomandibular - loses its suspension function and turns into a stapes - the first auditory ossicle located in the cavity of the middle ear, formed from the squirter. The lower element of the hyoid arch - the hyoid and part of the branchial arches form the hyoid oral plate with paired horns to support the muscles of the bottom of the oropharyngeal cavity and tongue.
The remains of the branchial arches, together with the hyoid, form the hyoid apparatus, and also form the cartilages of the larynx.
Paired limbs built like a system of levers connected by articulated joints. The forelimbs of tailless animals are much shorter than the hind limbs.
Forelimb is formed by the following elements:
1) Shoulder - humerus;
2) Forearm - forearm bone (fused ulna and radius bones);
a) wrist – 8–10 bones, arranged in 2–3 rows;
b) metacarpus – 4 elongated bones (according to the number of fingers), arranged in 1 row;
c) phalanges of the fingers - several short tubular bones that make up 4 fingers and the rudiment of the fore-finger.
Hind limb consists of the following elements:
1) Femur – femur;
2) Tibia – shin bone (fused tibia and fibula);
a) tarsus – two bones of the proximal row are elongated and form an additional lever, 2-3 bones are preserved from the distal row of the tarsus;
b) metatarsus – 5 elongated bones (according to the number of fingers), arranged in 1 row;
c) phalanges of the fingers - several short tubular bones, making up 5 fingers, connected by a swimming membrane.
Limb belts:
Forelimb belt (shoulder girdle) has the appearance of a semiring lying deep in the muscles. The main elements are paired scapulae and coracoids, forming articular sockets for articulation with the head of the humerus. Wide suprascapular cartilages are attached to the shoulder blades for attaching the back muscles, and cartilaginous procoracoids are attached to the coracoids, the lower side of which strengthens the clavicles. The inner ends of the procoracoids and coracoids of the right and left sides merge with each other. The bony sternum with a cartilaginous end extends posteriorly from the fusion site, and the presternalum, also with a cartilaginous end, extends anteriorly.
Hind limb belt (pelvic girdle) consists of 3 elements, at the junction of which the acetabulum is formed for attaching the femoral head. These are the long iliac bones that attach the pelvic girdle to the sacral vertebra of the axial skeleton, the ischial bones and the pubic cartilage.
Skeletal structure
The skull of amphibians has undergone noticeable changes compared to that of fish. It exhibits characteristics characteristic of typical terrestrial animals. The greatest transformations occur in the visceral skull, where autostyly appears (the palatoquadrate cartilage grows to the braincase) and secondary upper jaws arise from the integumentary bones. The hyoid arch is transformed into an element of the hearing aid (stapes) and the hyoid plate. The gill apparatus is reduced.
The brain skull has a small number of replacement and covering bones, and therefore it remains predominantly cartilaginous for life (Fig. 46).
The occipital region of the skull contains only two lateral occipital bones of chondral origin. Both of them carry condyles, with the help of which the skull is attached to the spine.
The sides of the skull in the area of the auditory capsules consist predominantly of cartilage. Here only two anterior auricular bones develop, lying laterally and slightly in front of the occipital bones. The anterior auricular bones are adjacent to the outer side of a pair of integumentary squamosal bones. In the area of the orbit in the anterior part, one ring-shaped sphenoid-olfactory bone develops. On the upper and lower sides it is covered by the frontoparietal bones and parasphenoid. The olfactory capsule remains entirely cartilaginous. All bones of the sides of the braincase, with the exception of the squamosal, are of chondral origin.
The roof of the skull is formed by the integumentary bones. The main part of the frog's braincase is covered by the frontoparietal bones, which arise as a result of the pairwise fusion of the frontal and parietal bones. In front of the frontoparietal bones, towards the end of the muzzle, there is a paired nasal bone.
The bottom of the skull is lined with bones of integumentary origin - a relatively large cruciform parasphenoid and a paired vomer lying in front of it. Frogs have vomer teeth on their vomers. Some bones of the visceral skull also take part in the formation of the bottom of the skull: in the area of the olfactory capsule, paired palatine bones adjoin the vomers, and two pterygoid bones underlie the orbit.
The visceral skull is represented by a number of arches.
The jaw arch of amphibians consists of cartilage and bone elements. The palatoquadrate cartilage (primary upper jaw) grows to the bottom of the frog's skull with its anterior and posterior ends (autostyly). On the lower surface of the palatoquadrate cartilage, the already mentioned integumentary palatine and pterygoid bones are formed. The quadrate bone does not develop, and the posterior part of the palatoquadrate cartilage remains cartilaginous.
The function of the upper jaws (“biting” jaws) is performed by the secondary jaws, consisting of the integumentary premaxillary, or premaxillary, and maxillary bones. These bones bear small teeth. The maxilla on each side articulates with the thin, rod-shaped quadratojugal bone.
The lower jaw is represented mainly by Meckel's cartilage, covered on the outside by paired integumentary dental and angular bones. The anterior end of Meckel's cartilage is transformed into the small geniomaxillary bone by chondral ossification.
Due to autostyly, the hyoid arch ceases to perform the function of attaching the jaws to the skull. In this regard, the hyomandibular, which served as a jaw suspension, turns into a small bone - a stirrup. The stapes is located in the middle ear cavity and functions as an auditory ossicle. The hyoids and copula form a cartilaginous hyoid plate, located between the branches of the lower jaw, and one pair of anterior horns (processes) extending from the plate.
The frog's spine consists of nine vertebrae and, unlike fish, consists of 4 sections: cervical, trunk, sacral and caudal (Fig. 47).
The cervical region is represented by a single vertebra, which differs from the trunk vertebrae in that it is devoid of transverse processes and its body is small.
The body section of tailless amphibians usually consists of seven vertebrae. The bodies of the trunk vertebrae have a concave surface in front and a convex surface in the back. Vertebrae of this structure are called prodel (Fig. 48). However, the last trunk vertebra has an amphicoelous type of structure. All vertebrae of the trunk are equipped with upper arches that form the spinal canal. Each arch ends in a weakly defined superior spinous process. Long transverse processes extend from the lateral sides of the vertebral bodies. At the base of the upper arches there are two pairs of articular processes: anterior and posterior.
The sacral section is represented by one vertebra. The bones of the hind limb girdle are attached to its transverse processes.
The caudal spine consists of a single bone called the urostyle. It is formed by the fusion of several caudal vertebrae, which are formed during the embryonic development of the frog.
The shoulder girdle (Fig. 49) has the form of an open ring or arc located in the front part of the torso. The dorsal part is represented by the scapula, to which the horizontally located suprascapular cartilage adjoins. Two other elements are located on the ventral side of the girdle: a relatively wide coracoid and an anterior cartilaginous procoracoid. On the latter lies the integumentary bone - the collarbone. The scapula, coracoid and procoracoid with the clavicle converge at the point of articulation of the forelimb, forming the articular fossa. The inner ends of the coracoid and procoracoid are bordered by procoracoid cartilage. Behind the suture between the procoracoid cartilages is a small sternum, and in front is the presternum. Both of these bones end in cartilage. Due to the absence of ribs, there is no chest, and the belt of the forelimbs lies freely in the thickness of the muscles.

Figure 49 – Shoulder girdle and forelimb of a frog
1 – shoulder blade; 2 – suprascapular cartilage; 3 – coracoid; 4 – procoracoid; 5 – collarbone; 6 – sternum; 7 – presternum; 8 – procoracoid cartilage;
9 – cartilaginous part of the sternum; 10 – cartilaginous part of the presternum; 11 – humerus; 12 – forearm (fused ulna and radius bones); 13 – carpal bones; 14 – metacarpal bones; 15 – phalanges of fingers.
The pelvic girdle (Fig. 50) of the frog, like the shoulder girdle, consists of three pairs of elements. The three elements of each side, by analogy with the shoulder girdle, are connected at the point of articulation of the limb and form here the articular fossa, or acetabulum.
The first pair of bones is called the ilia. Their function is to attach the hind limb girdle to the axial skeleton through the transverse processes of the sacral vertebra. In the frog, due to its peculiar method of movement - jumping, the iliae have elongated anterior ends - the wings of the ilium. The second pair of bones - the ischium - are located below and slightly behind the acetabulum. The third paired element - the pubis - remains cartilaginous in the frog. The pubic cartilages are directed forward and downward.

Figure 50 – Pelvic girdle and hind limb of a frog
1 – ilium; 2 – ischium; 3 – pubic cartilage;
4 – femur; 5 – lower leg (fused tibia and fibula); 6 – tarsal ring (fused upper tarsal bones); 7 – distal tarsal bones; 8 – metatarsal bones; 9 – rudiment of the 6th finger; 10 – phalanges of fingers.
Thus, the girdles of the frog's limbs are built according to a single pattern, common to both the shoulder and pelvic girdles. In general, this scheme is also typical for most terrestrial vertebrates. The appearance of limb girdles of this type and the emergence of their connection with the axial skeleton (pelvic girdle) also indicates the progressiveness of the structure of the amphibian skeleton in comparison with the skeleton of fish.
The fore and hind limbs of the frog are built according to a single pattern, characteristic of all terrestrial vertebrates. In a typical case, the five-fingered limb of a terrestrial vertebrate consists of three main sections, the last (distal) of which, in turn, splits into three subsections.
Section I – shoulder in the forelimb, thigh in the hindlimb. This section is always represented by one bone, which enters with its proximal end into the articular cavity of the corresponding girdle.
Section II – forearm in the forelimb, tibia in the hindlimb. The section consists of two bones: the ulna and radius in the forearm, and the tibia and fibula in the lower leg.
Section III – hand in the forelimb, foot in the hindlimb. This department has 3 subdivisions.
The structure of the frog's limbs is somewhat different from the above diagram, which is caused by a specific method of movement.
In the forelimb (Fig. 49), these differences are expressed in the fact that the ulna and radius bones following the shoulder fuse into one bone of the forearm. The number of fingers has been reduced to four.
In the hind limb (Fig. 50), the tibia is represented by only one bone, and not two, as follows from the diagram discussed above. The shin bone is formed as a result of the fusion of the tibia and fibula. The two proximal tarsal bones become greatly enlarged and, fused at their ends, form a kind of ring. The remaining tarsal bones either fuse or are reduced. In front of the first (inner) finger, a rudiment of an additional (sixth) finger is preserved.