Kas seotud pärandi teooria on tsütoloogiliselt kinnitatud? Seotud geenide pärand. Pärilikkuse kromosoomiteooria T. Morgan. Vaadake üle küsimused ja ülesanded
Seotud pärand - tunnuste pärand, mille geenid paiknevad samas kromosoomis. Geenidevahelise sideme tugevus oleneb nendevahelisest kaugusest: mida kaugemal geenid üksteisest asuvad, seda suurem on ristumise sagedus ja vastupidi. Sõltumatult päritavate tunnuste kõrval peavad olema ka pärilikud tunnused, mis on omavahel seotud, kuna need on määratud samas kromosoomis paiknevate geenide poolt. Sellised geenid moodustuvad sidurigrupp. Aheldusrühmade arv teatud liigi organismides võrdub kromosoomide arvuga haploidses komplektis (näiteks Drosophilal 1 paar = 4, inimestel 1 paar = 23).
Täielik haardumine- omamoodi seotud pärand, mille puhul analüüsitavate tunnuste geenid paiknevad üksteisele nii lähedal, et nendevaheline ristumine muutub võimatuks.
Mittekomplektne sidur- seotud pärilikkuse tüüp, mille puhul analüüsitavate tunnuste geenid asuvad üksteisest teatud kaugusel, mis teeb võimalikuks nendevahelise ristumise.
(Ristsugurakud- sugurakud, mille moodustumise käigus toimus ristumine. Ristsugurakud moodustavad reeglina väikese osa sugurakkude koguarvust.
Üleminek- homoloogsete kromosoomide sektsioonide vahetus raku jagunemise protsessis, peamiselt esimese meiootilise jagunemise profaasis, mõnikord mitoosis. T. Morgani, K. Bridgesi ja A. Sturtevandi katsed näitasid, et geenide absoluutselt täielikku seost, mille puhul geenid alati koos edasi kanduks, ei eksisteeri. Tõenäosus, et kaks samas kromosoomis asuvat geeni ei lahkne meioosi ajal, on vahemikus 1-0,5. Looduses domineerib mittetäielik aheldus homoloogsete kromosoomide ristumise ja geenide rekombinatsiooni tõttu. Ülemineku tsütoloogilist mustrit kirjeldas esmakordselt Taani teadlane F. Janssens.
Üleminek toimub ainult siis, kui geenid on heterosügootses olekus (AB / av). Kui geenid on homosügootses olekus (AB / AB või aB / aB), ei anna identsete piirkondade vahetus sugurakkudes ja põlvkonnas uusi geenide kombinatsioone. Geenidevahelise ristumise sagedus (protsent) sõltub nendevahelisest kaugusest: mida kaugemal nad üksteisest asuvad, seda sagedamini toimub ristumine. T. Morgan tegi ettepaneku mõõta geenidevahelist kaugust ristumisega protsendina valemi järgi:
N1/N2 X 100 = Crossover %,
kus N1 on isendite koguarv F-s;
N2 on ristuvate indiviidide koguarv.
Kromosoomi segment, millel toimub 1% ristumisest, on võrdne ühe morganiidiga (geenidevahelise kauguse tingimuslik mõõt). Ületamise sagedust kasutatakse geenide suhtelise asukoha ja nendevahelise kauguse määramiseks. Inimese geenikaardi koostamiseks kasutatakse uusi tehnoloogiaid, lisaks on ehitatud kromosoomide tsütogeneetilisi kaarte.
Ületusi on mitut tüüpi: kahekordne, mitmekordne (keeruline), ebakorrapärane, ebaühtlane.
Üleminek toob kaasa uue geenide kombinatsiooni, põhjustades fenotüübi muutuse. Lisaks on see koos mutatsioonidega oluline tegur organismide evolutsioonis.)
T. Morgani uurimistöö tulemus oli tema pärilikkuse kromosoomiteooria loomine:
· geenid paiknevad kromosoomides; erinevad kromosoomid sisaldavad ebavõrdset arvu geene; iga mittehomoloogse kromosoomi geenide komplekt on ainulaadne;
· igal geenil on kromosoomis kindel asukoht (lookus); alleelgeenid paiknevad homoloogsete kromosoomide identsetes lookustes;
· geenid paiknevad kromosoomidel kindlas lineaarses järjestuses;
· samas kromosoomis asuvad geenid päranduvad koos, moodustades aheldusrühma; aheldusrühmade arv on võrdne haploidse kromosoomide komplektiga ja on iga organismitüübi puhul konstantne;
· geeniside võib katkeda ristumise käigus, mis viib rekombinantsete kromosoomide moodustumiseni; ületamise sagedus sõltub geenidevahelisest kaugusest: mida suurem vahemaa, seda suurem on ületamise väärtus;
· igal liigil on ainult talle iseloomulik kromosoomide komplekt – karüotüüp.
Soo ja sooga seotud tunnuste pärand. Sugukromosoomid ja nende roll soo määramisel. Seksuaalne pärand. Isendi sugu on keeruline tunnus, mis kujuneb nii geenide toimel kui ka arengutingimustel. Inimestel on üheks 23-st kromosoomipaarist sugukromosoomid, mida tähistatakse kui X ja Y. Naised on homogameetiline sugu, st. Neil on kaks X-kromosoomi, üks nende emalt ja teine isalt. Isased on heterogameetilised, neil on üks X-üks Y-kromosoom ja X edastatakse emalt ja Y isalt. Pange tähele, et heterogameetiline sugu ei pruugi alati olla mees; näiteks lindudel on need emased, isased aga homogameetilised. On ka teisi soo määramise mehhanisme. Seega puudub paljudel putukatel Y-kromosoom. Sel juhul areneb üks soost kahe X-kromosoomi ja teine - ühe X-kromosoomi juuresolekul. Mõnel putukatel määrab sugu autosoomide ja sugukromosoomide arvu suhe. Paljudel loomadel on nn. soo ümbermääratlemine, kui sigoot areneb olenevalt keskkonnateguritest kas emaseks või isaseks. Taimede soo arengul on sama mitmekesised geneetilised mehhanismid kui loomadel.
X-kromosoomiga seotud tunnused. Kui geen asub sugukromosoomis (seda nimetatakse suguga seotud), siis järgib selle ekspressioon järglastel muid reegleid kui autosomaalsete geenide puhul. Mõelge geenidele, mis asuvad X-kromosoomis. Tütar pärib kaks X-kromosoomi, ühe emalt ja teise isalt. Pojal on ainult üks X-kromosoom – emalt; isalt saab ta Y-kromosoomi. Seetõttu annab isa oma X-kromosoomis olevad geenid edasi ainult tütrele, samas kui poeg ei saa neid vastu võtta. Kuna X-kromosoom on Y-kromosoomiga võrreldes geenide poolest "rikkam", siis selles mõttes on tütar geneetiliselt sarnasem isaga kui poeg; poeg on rohkem nagu oma ema kui isa.
Üks ajalooliselt tuntumaid sooga seotud tunnuseid inimestel on hemofiilia, mille tagajärjeks on tõsine verejooks väikestest sisselõigetest ja ulatuslikud verevalumid verevalumitest. Seda põhjustab retsessiivne defektne alleel 0, mis blokeerib vere hüübimiseks vajaliku valgu sünteesi. Selle valgu geen asub X-kromosoomis. Heterosügootsel naisel +0 (+ tähendab normaalset aktiivset alleeli, mis domineerib hemofiilia 0 alleeli üle) ei arene hemofiilia ega ka tema tütrel, kui isal seda patoloogiat pole. Tema poeg võib aga saada 0-alleeli ja seejärel areneb tal hemofiilia. X-kromosoomi geenidest põhjustatud retsessiivsed haigused mõjutavad naisi palju harvemini kui mehi, kuna neil on haigus ainult siis, kui nad on homosügootsed - retsessiivse alleeli olemasolu mõlemas homoloogses X-kromosoomis; mehed haigestuvad alati, kui nende üksik X-kromosoom kannab defektset alleeli.
Seos Y-kromosoomiga.Infot Y-kromosoomis paiknevate geenide kohta on väga vähe. Eeldatakse, et see praktiliselt ei kanna geene, mis määravad raku toimimiseks vajalike valkude sünteesi. Kuid see mängib võtmerolli meessoost fenotüübi kujunemisel. Y-kromosoomi puudumine ainult ühe X-kromosoomi olemasolul viib nn. Turneri sündroom: nõrgalt arenenud esmaste ja sekundaarsete seksuaalomaduste ning muude kõrvalekalletega naissoost fenotüübi areng. On mehi, kellel on täiendav Y-kromosoom (XYY); nad on pikad, agressiivsed ja sageli ebatavalise käitumisega. Y-kromosoomis on tuvastatud mitu geeni, mis vastutavad spetsiifiliste ensüümide ja hormoonide sünteesi reguleerimise eest ning nende rikkumised põhjustavad seksuaalse arengu patoloogiaid. On mitmeid morfoloogilisi tunnuseid, mis arvatakse olevat määratud Y-kromosoomi geenidega; nende hulgas on kõrvakarvade areng. Sellised märgid edastatakse ainult meesliini kaudu: isalt pojale.
geneetiline soo määramine, määratud sugukromosoomide komplektiga, toetab emaste ja isaste võrdset paljunemist. Tõepoolest, emaste munad sisaldavad ainult X-kromosoomi, kuna naistel on sugukromosoomides XX genotüüp. Meeste genotüüp on XY ja seetõttu määrab tüdruku või poisi sünni igal juhul see, kas sperma kannab X- või Y-kromosoomi. Kuna meioosi protsessis on kromosoomidel võrdne võimalus sugurakku sattuda, siis pooled isasloomade poolt toodetud sugurakkudest sisaldavad X-kromosoomi ja pooled Y-kromosoomi. Seetõttu eeldatakse, et pooled järglastest on ühest ja pooled teisest soost.
Tuleb rõhutada, et poisi või tüdruku sündi on võimatu ette ennustada, kuna on võimatu ennustada, milline meessoost sugurakk osaleb munaraku viljastumises: see, mis kannab X- või Y-kromosoomi. . Seetõttu on enam-vähem poiste olemasolu peres juhuse küsimus.
Seotud pärilikkuse nähtuste analüüs, ristumine, geneetiliste ja tsütoloogiliste kaartide võrdlemine võimaldavad sõnastada pärilikkuse kromosoomiteooria põhisätted:
Geenid asuvad kromosoomides. Lisaks sisaldavad erinevad kromosoomid ebavõrdset arvu geene. Lisaks on iga mittehomoloogse kromosoomi geenide komplekt ainulaadne.
Alleelsed geenid hõivavad sama loci homoloogsetel kromosoomidel.
Geenid paiknevad kromosoomis lineaarses järjestuses.
Ühe kromosoomi geenid moodustavad aheldusrühma ehk päranduvad valdavalt seotuna (ühiselt), mille tõttu tekib mõne tunnuse seotud pärand. Aheldusrühmade arv on võrdne antud liigi kromosoomide haploidse arvuga (homogameetilises soos) või rohkem kui 1 (heterogameetilises soos).
Aheldus katkeb ristumise tulemusena, mille sagedus on otseselt võrdeline kromosoomi geenide vahelise kaugusega (seetõttu on sideme tugevus pöördvõrdeline geenidevahelise kaugusega).
Igat bioloogilist liiki iseloomustab teatud kromosoomide komplekt - karüotüüp.
Seotud pärand
Sõltumatu funktsioonide kombinatsioon ( Mendeli kolmas seadus) viiakse läbi tingimusel, et neid tunnuseid määravad geenid asuvad erinevates homoloogsete kromosoomide paarides. Seetõttu on igas organismis meioosis iseseisvalt kombineeritud geenide arv piiratud kromosoomide arvuga. Kuid organismis ületab geenide arv oluliselt kromosoomide arvu. Näiteks enne molekulaarbioloogia ajastut uuriti maisil rohkem kui 500 geeni, kärbsel Drosophila üle 1 tuhande ja inimestel umbes 2 tuhat geeni, samal ajal kui neil on vastavalt 10, 4 ja 23 paari kromosoome. See, et geenide arv kõrgemates organismides on mitu tuhat, oli W. Settonile selge juba 20. sajandi alguses. See andis põhjust oletada, et igas kromosoomis on lokaliseeritud palju geene. Samas kromosoomis asuvad geenid moodustavad aheldusrühma ja on päritud koos.
Ühine geenide pärand T. Morgan tegi ettepaneku nimetada seotud pärand. Aheldusrühmade arv vastab kromosoomide haploidsele arvule, kuna aheldusrühm koosneb kahest homoloogsest kromosoomist, milles paiknevad samad geenid. (Heterogameetilisest soost isenditel, näiteks isastel imetajatel, on tegelikult veel üks sidestusrühm, kuna X- ja Y-kromosoomid sisaldavad erinevaid geene ja esindavad kahte erinevat aheldusrühma. Seega on naistel 23 siderühma ja meestel - 24).
Seotud geenide pärandumise viis erineb homoloogsete kromosoomide erinevates paarides paiknevate geenide pärandusest. Seega, kui sõltumatu kombinatsiooniga moodustab diheterosügootne isend võrdsetes kogustes nelja tüüpi sugurakke (AB, Ab, aB ja ab), siis seotud pärandiga (ületamise puudumisel) moodustab sama diheterosügoot ainult kahte tüüpi sugurakke. sugurakud: (AB ja ab) ka võrdsetes kogustes. Viimased kordavad vanema kromosoomi geenide kombinatsiooni.
Leiti aga, et uute geenikombinatsioonidega - Ab ja aB - tekivad lisaks tavalistele (mitte-ristuvate) sugurakkudele ka teised (ristsugurakud) sugurakud, mis erinevad vanema kromosoomide geenide kombinatsioonidest. Selliste sugurakkude tekke põhjuseks on homoloogsete kromosoomide lõikude vahetus ehk üle minemine.
Üleminek toimub meioosi I profaasis ajal konjugatsioonid homoloogsed kromosoomid. Sel ajal võivad kahe kromosoomi osad ristuda ja oma osi vahetada. Selle tulemusena tekivad kvalitatiivselt uued kromosoomid, mis sisaldavad nii ema kui ka isa kromosoomide sektsioone (geene). Isikuid, kes on saadud sellistest sugurakkudest uue alleelide kombinatsiooniga, nimetatakse ristuvateks või rekombinantseteks.
Ristumissagedus (protsent) kahe samas kromosoomis asuva geeni vahel on võrdeline nendevahelise kaugusega. Kahe geeni vaheline ristumine toimub seda harvemini, mida lähemal need üksteisele on. Kui geenide vaheline kaugus suureneb, suureneb tõenäosus, et üleminek eraldab need kahel erineval homoloogsel kromosoomil.
Geenidevaheline kaugus iseloomustab nende sideme tugevust. On geene, mille ahelduse protsent on suur, ja neid, kus seost peaaegu ei tuvastata. Seotud pärimise korral ei ületa maksimaalne ristumise sagedus aga 50%. Kui see on kõrgem, siis on alleelipaaride vahel vaba kombinatsioon, mida ei saa eristada sõltumatust pärandist.
Ületamise bioloogiline tähtsus on äärmiselt suur, kuna geneetiline rekombinatsioon võimaldab luua uusi, varem olematuid geenikombinatsioone ja seeläbi suurendada pärilikkust varieeruvus, mis annab rohkelt võimalusi organismi kohanemiseks erinevates keskkonnatingimustes. Spetsiaalselt dirigeerib inimene hübridisatsioon aastal kasutamiseks vajalike kombinatsioonide saamiseks aretustöö.
Üleminek. See protsess toimub meioosi I profaasis ajal, mil homoloogsed kromosoomid on konjugatsiooni tulemusena tihedalt kokku viidud ja moodustavad kahevalentseid. Ristumise käigus vahetatakse vastavaid sektsioone homoloogsete kromosoomide vastastikku põimuvate kromatiidide vahel (joonis 3.72). See protsess tagab igas aheldusrühmas geenide isa- ja emaalleelide rekombinatsiooni. Sugurakkude erinevates prekursorites toimub ristumine kromosoomide erinevates osades, mille tulemuseks on kromosoomides mitmesugused vanemate alleelide kombinatsioonid.
Riis. 3.72. Ristumine sugurakkude geneetilise mitmekesisuse allikana:
I - vanemate sugurakkude viljastamine a ja b c sigootide moodustumine sisse;II - gametogenees sügoodist arenenud organismis sisse;G- homoloogide vaheline üleminek profaasis ma; d - rakud, mis moodustuvad pärast 1. meiootilist jagunemist; e, w - rakud, mis moodustuvad pärast meioosi teist jagunemist ( e - mitte-ristuvad sugurakud, millel on algsed vanemlikud kromosoomid; hästi - ristuvad sugurakud koos päriliku materjali rekombinatsiooniga homoloogsetes kromosoomides)
On selge, et ristumine kui rekombinatsioonimehhanism on efektiivne ainult siis, kui isa ja ema kromosoomide vastavad geenid on esindatud erinevate alleelidega. Absoluutselt identsed aheldusrühmad ristumise ajal ei anna uusi alleelide kombinatsioone.
Üleminek ei toimu meioosi ajal mitte ainult sugurakkude prekursorites. Seda täheldatakse ka somaatilistes rakkudes mitoosi ajal. Somaatilist ristumist on kirjeldatud Drosophila ja mõne hallituse liigi puhul. See viiakse läbi homoloogsete kromosoomide vahelise mitoosi ajal, kuid selle sagedus on 10 000 korda väiksem kui meiootilise ristumise sagedus, mille mehhanismist see ei erine. Mitootilise ristumise tulemusena tekivad somaatiliste rakkude kloonid, mis erinevad neis üksikute geenialleelide sisalduse poolest. Kui sügooti genotüübis on see geen esindatud kahe erineva alleeliga, siis somaatilise ristumise tulemusena võivad tekkida selle geeni samade isa- või emaalleelidega rakud (joonis 3.73).

Riis. 3.73. Üleminek somaatilistes rakkudes:
1 - somaatiline rakk, mille homoloogsetes kromosoomides on geen A esindatud kahe erineva alleeliga (A ja a); 2 - ülesõit; 3 - homoloogsete kromosoomide vahelise vastavate lõikude vahetuse tulemus; 4 - homoloogide paiknemine lõhustumisspindli ekvatoriaaltasandil mitoosi metafaasis (kaks varianti); 5 - tütarrakkude moodustumine; 6 - geeni A suhtes heterosügootsete rakkude moodustumine, mis on alleelide komplekti (Aa) poolest sarnased emarakuga; 7 - geeni A suhtes homosügootsete rakkude moodustumine, mis erinevad emarakust alleelide komplekti (AA või aa) poolest
19. sajandi keskel, kui G. Mendel viis läbi oma katseid ja sõnastas geneetika ja laiemalt bioloogia arengule universaalse ja fundamentaalse tähtsusega mustreid, ei piisanud veel teaduslikest teadmistest, et mõista pärimismehhanisme. Seetõttu oli Mendeli tööd aastaid tagasi nõudmata. Kuid 20. sajandi alguseks. olukord bioloogias on radikaalselt muutunud.
Avastati mitoos ja meioos, taasavastati Mendeli seadused. Üksteisest sõltumatult tegid Saksamaa ja USA teadlased ettepaneku, et pärilikud tegurid paiknevad kromosoomides. 1906. aastal kirjeldas R. Pennet esimest korda kahe tunnuse iseseisva pärimise Mendeli seaduse rikkumist. Klassikalise õievärvi ja õietolmu kuju poolest erinevate magushernetaimede dihübriidristi lavastamisel ei saanud Pennet teises põlvkonnas oodatud lõhenemist 9:3:3:1. F2 hübriididel olid ainult vanemlikud fenotüübid vahekorras 3:1, st tunnuste ümberjaotumist ei toimunud.
Tasapisi kogunes üha rohkem selliseid erandeid, mis ei allunud iseseisva pärimise seadusele. Tekkis küsimus, kuidas täpselt geenid kromosoomides paiknevad? Lõppude lõpuks on tunnuste arv ja seega ka geenide arv igas organismis palju suurem kui kromosoomide arv. See tähendab, et iga kromosoom sisaldab palju geene, mis vastutavad erinevate tunnuste eest. Kuidas päritakse samas kromosoomis asuvad geenid?
■ T. Morgani töö. Nendele küsimustele vastas rühm Ameerika teadlasi eesotsas Thomas Hunt Morganiga (1866-1945). Töötades väga mugava geneetilise objekti – äädikakärbse Drosophila kallal, said nad suurepäraselt hakkama geenide pärilikkuse uurimisega.
Teadlased on selle leidnud geenid, mis asuvad samas kromosoomis, on päritud koos, st seotud. Sellele nähtusele on antud nimi morgani seadus, või seotud pärimise seadus . Ühes kromosoomis paiknevaid geenirühmi nimetatakse sidurigrupp. Kuna homoloogsetes kromosoomides asuvad identsed geenid, võrdub aheldusrühmade arv kromosoomipaaride arvuga, st kromosoomide haploidse arvuga. Inimesel on 23 paari kromosoome ja seega 23 siderühma, koeral 39 paari kromosoome ja 39 siderühma, hernel 7 paari kromosoome ja 7 siderühma jne. Tuleb märkida, et Mendelil vedas üllatavalt dihübriidristide loomisel: erinevate tunnuste (herneste värvus ja kuju) eest vastutavad geenid paiknesid erinevates kromosoomides. Oleks võinud olla teisiti ja siis poleks iseseisva lõhenemise mustrit avastanud.
T. Morgani rühma töö tulemuseks oli 1911. a pärilikkuse kromosoomiteooria.
Mõelge tänapäevase pärilikkuse kromosoomiteooria põhisätetele.
Pärilikkuse ühik on geen, mis on kromosoomi segment.
Geenid paiknevad kromosoomidel rangelt määratletud kohtades (lookuses) ja alleelsed geenid (vastutavad ühe tunnuse kujunemise eest) paiknevad homoloogsete kromosoomide samades lookustes.
Geenid paiknevad kromosoomides lineaarses järjekorras, st üksteise järel.
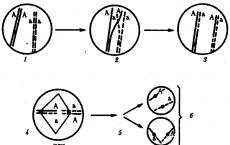
■Siduri rike. Samas kromosoomis paiknevate geenide pärilikkuse analüüsimisel leiti mõnes ristamises aga seostushäire. Selgus, et mõnikord võivad paaris olevad homoloogsed kromosoomid omavahel identseid homoloogseid piirkondi vahetada. Et see juhtuks, peavad kromosoomid asuma üksteise vahetus läheduses. Seda homoloogsete kromosoomide ajutist paaripõhist lähenemist nimetatakse konjugatsiooniks. Sel juhul võivad kromosoomid vahetada üksteise vastas paiknevaid lookusi, mis sisaldavad samu geene. Seda nähtust nimetatakse ületamiseks.
Tuletage meelde meioosi jagunemist, mille käigus moodustuvad sugurakud. Esimese meiootilise jagunemise profaasis võib bivalentse (tetradi) moodustumisel, kui kahekordistunud homoloogsed kromosoomid seisavad üksteisega paralleelselt, võib toimuda sarnane vahetus. Selline sündmus viib geneetilise materjali rekombinatsioonini, suurendab järeltulijate mitmekesisust, st suurendab pärilikku varieeruvust ja mängib seetõttu evolutsioonis olulist rolli.
■ Geneetilised kaardid. Alleelsete geenide vahetuse nähtus homoloogsete kromosoomide vahel aitas teadlastel kindlaks teha iga geeni asukoha kromosoomis, st koostada geneetilisi kaarte. Kromosoomi geneetiline kaart on diagramm geenide vastastikusest paigutusest, mis asuvad samas kromosoomis, st samas aheldusrühmas. Selliste kaartide koostamine pakub suurt huvi nii fundamentaaluuringute kui ka mitmesuguste praktiliste probleemide lahendamiseks. Näiteks inimese kromosoomide geneetilised kaardid on väga olulised mitmete raskete pärilike haiguste diagnoosimisel.
Praegu asendatakse lihtsad geneetilised kaardid molekulaargeneetiliste kaartidega, mis sisaldavad teavet geenide nukleotiidjärjestuste kohta.
Küsimused enesekontrolliks
1. Mis on seotud pärimine?
2. Mis on geenide aheldusrühmad?
3. Mis on geenisideme häire põhjus?
4. Mis on homoloogsete kromosoomide vahelise alleelsete geenide vahetuse bioloogiline tähtsus?
5. Kas seotud pärandi teooria on tsütoloogiliselt kinnitatud?
G. Mendeli järgi päranduvad geenid iseseisvalt ja paiknevad erinevates kromosoomides. Diheterosügoot AaBb tsütoloogilise valemi kujul võib esitada järgmiselt:
Meioosi ajal moodustab see 4 tüüpi sugurakke: AB, Ab, aB, ab.
Aastatel 1911–1912 Ameerika geneetik T. Morgan tõestas, et samas kromosoomis asuvad geenid võivad olla pärilikult seotud (koos). See on tingitud asjaolust, et geene on palju rohkem kui kromosoome. Geenide kaaspärimine piirates nende vaba kombinatsiooni, helistas T. Morgan geenide seos või seotud pärand. Kui geenid on päritud seotud, võib AaBb diheterosügoodi genotüüpi tsütoloogilise valemi abil esitada järgmiselt:
Meioosi ajal, kui geenid on täielikult seotud, moodustab see 2 tüüpi sugurakke - AB, ab. Selliseid sugurakke nimetatakse mitte-ristuvateks.
T. Morgani ja tema õpilaste uuringud näitasid, et geene vahetatakse regulaarselt homoloogses kromosoomipaaris. Protsessi, mille käigus vahetatakse homoloogsete kromosoomide identsed lõigud neis paiknevate geenidega, nimetatakse kromosoomide decussatsiooniks või üle minemine.Üleminek annab uued geenide kombinatsioonid, mis asuvad homoloogsetel kromosoomidel . Seetõttu on ülesõidu fenomenist tulenevalt lisaks mitte-crossoverile ka ristuvad sugurakud - Ab, aB.
T. Morgani seadus on sõnastatud järgmiselt: samas kromosoomis asuvad geenid on päritud seotud (koos) ja seose tugevus sõltub geenide vahelisest kaugusest.
T. Morgani kromosoomiteooria põhisätted:
1) geenid paiknevad kromosoomidel ja erinevates kromosoomides on ebavõrdne arv geene. Iga mittehomoloogse kromosoomi geenide komplekt on ainulaadne;
2) geenid paiknevad kromosoomis kindlas lineaarses järjestuses;
3) geenid moodustavad aheldusrühmi ja võivad olla pärilikult seotud. Aheldusrühmade arv vastab haploidsele kromosoomide komplektile (inimesel on 23 siderühma);
4) homoloogsete kromosoomide vahel on võimalik ristumine (crossover) ja alleelgeenide vahetus ning ristumine toimub siis, kui geenide vahel on vahemaa;
5) geenide vaheline kaugus kromosoomis on võrdeline ristumise sagedusega ja väljendatakse ristumise protsendina.
Geenidevahelise ristumise protsent arvutatakse järgmise valemiga:
X= ––––
a on ühe klassi ristuvate indiviidide arv; b on teise klassi ristuvate indiviidide arv;
n on isendite koguarv; Х – geenidevahelise ristumise % (geenidevaheline kaugus ristumisprotsentides).
Geenide vahelist kaugust mõõdetakse spetsiaalsetes ühikutes - morganiidid. 1 morganiid (1 M) = 1% ristumist.
Geeniuuringutes kasutati äädikakärbest, millel on 8 kromosoomist koosnev karüotüüp, viljakus ja palju alternatiivseid tunnuseid. T. Morgan käsitles olukorda, mil geenide täielik aheldus ja geenide ahelduse rikkumine (üleminek) esinevad inimesel samal ajal. Esimeses etapis võeti ristamise jaoks homosügootsed isendid (P ♀AABB x ♂aabb). Selle tulemusena olid esimeses põlvkonnas kõik isendid diheterosügootsed (AaBb).
Geeni tunnus
Hall kehavärvi
ja tume kehavärv
Pikkades tiibades
b algelised tiivad.
Täielik geenide seos
P ♀ ══x ♂ ══
G AB, ab ab - meessugurakud
(Naiste sugurakud, mis ei ole ristuvad)
F 1 ══ ; ══
Geenide täieliku ahelduse korral täheldatakse 1:1 jagunemist. Kokku saadi 83% isenditest, kes pärisid oma vanemate fenotüübi, kuna nad moodustati mitte-ristuvatest sugurakkudest. Nende hulgas oli halli kehaga, normaalsete tiibadega isendeid 41,5% ja musta kehaga, algeliste tiibadega isendeid samuti 41,5%.
Geenisideme häire
P ♀ ══ x ♂ ══
G Ab, aB ab - meessugurakud
(Ristuvad naiste sugurakud)
F 1 ══; ══
Geenide mittetäieliku ahelduse korral täheldatakse ka 1:1 lõhenemist. Kokku saadi 17% rekombinantsete fenotüüpidega indiviididest, kuna need moodustati ristuvatest sugurakkudest. Isikuid, kellel on uus tunnuste kombinatsioon, nimetatakse rekombinantsed. Rekombinantsetest moodustasid halli kehaga kärbsed, algelised tiivad 8,5% ja musta kehaga normaalsed tiivad samuti 8,5%.
Küsimus 1. Mis on seotud pärimine?
Seotud pärand on ühes kromosoomis (st ühes DNA molekulis) paiknevate geenide ühine pärand. Näiteks magusatel hernestel on niimoodi paigutatud õite värvust ja õietolmu kuju määravad geenid. Need päranduvad lingitud viisil, seetõttu moodustuvad teise põlvkonna hübriidide ristamisel vanemlikud fenotüübid vahekorras 3:1 ja iseseisva pärandusega dihübriidsele ristumisele iseloomulik lõhenemine 9:3:3:1. ei ilmu.
Küsimus 2. Mis on geenide aheldusrühmad?
Aheldusrühm on geenide kogum, mis asub samas kromosoomis. Kuna homoloogsed kromosoomid sisaldavad samu geene, võrdub sidestusrühmade arv kromosoomide haploidse arvuga (inimestel 23, hernestel 7, Drosophilal 4).
3. küsimus. Mis on geenisideme häire põhjus?
Geenisideme katkemise põhjuseks on homoloogsete kromosoomide lõikude vahetus meiootilise jagunemise I profaasis. Tuletame meelde, et selles etapis konjugeeritakse paariskromosoomid, moodustades niinimetatud kahevalentsed ribad. Bivalentide moodustumine võib viia kromosoomide ristumiseni, mis loob võimaluse homoloogsete DNA piirkondade vahetamiseks. Kui see juhtub, siis aheldusrühmad muudavad oma sisu (sisaldavad samade geenide teisi alleele) ja järglaste hulka võivad ilmuda isendid, kelle fenotüüp erineb vanemate omadest.
Küsimus 4. Mis on homoloogsete kromosoomide vahelise alleelgeenide vahetuse bioloogiline tähtsus?materjali saidilt
Kromosoomide ristumine (muidu - üleminek) toob kaasa geneetilise materjali rekombinatsiooni ja uute aheldusrühma geenide alleelide kombinatsioonide moodustumise. Samal ajal suureneb järglaste mitmekesisus, s.t suureneb pärilik varieeruvus, millel on suur evolutsiooniline tähtsus. Tõepoolest, kui näiteks Drosophilas asuvad keha värvi ja tiibade pikkuse määravad geenid samas kromosoomis, siis normaalsete tiibadega hallide ja lühendatud tiibadega mustade kärbeste puhtaid jooni ristades, ristumise puudumise tõttu ei saa me kunagi muid fenotüüpe. Kromosoomide ristumise olemasolu võimaldab (mõnel protsendil juhtudest) ilmuda lühikeste tiibadega hall-kärbseid ja normaalsete tiibadega musti kärbseid.
Küsimus 5. Kas seotud pärilikkuse teooria on tsütoloogiliselt kinnitatud?
Thomas Hunt Morgani (1866-1945) teooriat seotud pärimise kohta kinnitavad tsütoloogilised vaatlused. Näidati, et jagunemise ajal lahknevad kromosoomid täielikult raku erinevatele poolustele. Järelikult jagunevad meioosi ajal samas kromosoomis asuvad geenid ühte sugurakku, see tähendab, et nad on tõesti päritud omavahel seotud viisil.
Kas te ei leidnud seda, mida otsisite? Kasutage otsingut