Vai saistītā mantojuma teorija ir apstiprināta citoloģiski? Saistītā gēnu pārmantošana. Iedzimtības hromosomu teorija T. Morgans. Pārskatiet jautājumus un uzdevumus
Saistīts mantojums - tādu pazīmju pārmantošana, kuru gēni ir lokalizēti vienā hromosomā. Saiknes stiprums starp gēniem ir atkarīgs no attāluma starp tiem: jo tālāk gēni atrodas viens no otra, jo lielāks ir krustošanās biežums un otrādi. Līdzās pazīmēm, kas tiek mantotas neatkarīgi, ir jābūt arī tām, kas tiek mantotas savstarpēji saistītas, jo tās nosaka gēni, kas atrodas vienā un tajā pašā hromosomā. Tādi gēni veidojas sajūga grupa. Saiknes grupu skaits noteiktas sugas organismos ir vienāds ar hromosomu skaitu haploīdajā komplektā (piemēram, Drosophila 1 pāris = 4, cilvēkiem 1 pāris = 23).
Pilna saķere- sava veida saistīta pārmantošana, kurā analizējamo pazīmju gēni atrodas tik tuvu viens otram, ka krustošanās starp tiem kļūst neiespējama.
Nepilnīgs sajūgs- Saistītā mantojuma veids, kurā analizējamo pazīmju gēni atrodas noteiktā attālumā viens no otra, kas ļauj to krustoties.
(Crossover gametas- gametas, kuru veidošanās procesā notika šķērsošana. Parasti krustojošās gametas veido nelielu daļu no kopējā gametu skaita.
Šķērsojot- homologu hromosomu sekciju apmaiņa šūnu dalīšanās procesā, galvenokārt pirmās meiotiskās dalīšanās profāzē, dažreiz mitozē. T. Morgana, K. Bridžesa un A. Stērtevanta eksperimenti parādīja, ka nepastāv absolūti pilnīga gēnu saite, kurā gēni vienmēr tiktu pārnesti kopā. Varbūtība, ka divi gēni, kas atrodas vienā hromosomā, meiozes laikā neatšķirsies, svārstās no 1 līdz 0,5. Dabā dominē nepilnīga saikne homologu hromosomu krustošanās un gēnu rekombinācijas dēļ. Citoloģisko krustošanās modeli pirmais aprakstīja dāņu zinātnieks F. Jansens.
Šķērsošana notiek tikai tad, ja gēni ir heterozigotā stāvoklī (AB / av). Ja gēni ir homozigotā stāvoklī (AB / AB vai aB / aB), identisku reģionu apmaiņa nedod jaunas gēnu kombinācijas gametās un paaudzē. Gēnu krustošanās biežums (procentos) ir atkarīgs no attāluma starp tiem: jo tālāk tie atrodas viens no otra, jo biežāk notiek krustošanās. T. Morgans ierosināja izmērīt attālumu starp gēniem, krustojot to procentos, pēc formulas:
N1/N2 X 100 = % Crossover,
kur N1 ir kopējais īpatņu skaits F;
N2 ir krosoveru personu kopējais skaits.
Hromosomas segments, uz kura notiek 1% krustošanās, ir vienāds ar vienu morganīdu (nosacīts attāluma mērījums starp gēniem). Šķērsošanas biežumu izmanto, lai noteiktu gēnu relatīvo stāvokli un attālumu starp tiem. Cilvēka ģenētiskās kartes izveidošanai tiek izmantotas jaunas tehnoloģijas, turklāt ir uzbūvētas hromosomu citoģenētiskās kartes.
Ir vairāki šķērsošanas veidi: dubultā, daudzkārtēja (sarežģīta), neregulāra, nevienmērīga.
Šķērsošana noved pie jaunas gēnu kombinācijas, izraisot izmaiņas fenotipā. Turklāt kopā ar mutācijām tas ir svarīgs faktors organismu evolūcijā.)
T. Morgana pētījuma rezultāts bija viņa radīja iedzimtības hromosomu teoriju:
· gēni atrodas hromosomās; dažādas hromosomas satur nevienlīdzīgu gēnu skaitu; gēnu kopums katrai nehomologajai hromosomai ir unikāls;
· katram gēnam ir noteikta vieta (lokuss) hromosomā; alēlie gēni atrodas identiskos homologo hromosomu lokusos;
· gēni atrodas hromosomās noteiktā lineārā secībā;
· gēni, kas atrodas vienā hromosomā, tiek mantoti kopā, veidojot saiknes grupu; saišu grupu skaits ir vienāds ar haploīdu hromosomu kopu un ir nemainīgs katram organisma tipam;
· gēnu saite var tikt pārtraukta šķērsošanas procesā, kas noved pie rekombinanto hromosomu veidošanās; šķērsošanas biežums ir atkarīgs no attāluma starp gēniem: jo lielāks attālums, jo lielāka ir šķērsošanas vērtība;
· katrai sugai ir tikai tai raksturīga hromosomu kopa - kariotips.
Dzimuma un ar dzimumu saistītu iezīmju pārmantošana. Dzimuma hromosomas un to nozīme dzimuma noteikšanā. Dzimuma pārmantošana. Indivīda dzimums ir sarežģīta iezīme, ko veido gan gēnu darbība, gan attīstības apstākļi. Cilvēkiem viens no 23 hromosomu pāriem ir dzimuma hromosomas, kas apzīmētas kā X un Y. Sievietes ir homogamētiskais dzimums, t.i. Viņiem ir divas X hromosomas, viena no mātes un viena no tēva. Tēviņi ir heterogamētiski, tiem ir viena X-viena Y-hromosoma, un X tiek pārnests no mātes, bet Y no tēva. Ņemiet vērā, ka heterogamētiskais dzimums ne vienmēr ir vīrietis; piemēram, putniem tās ir mātītes, savukārt tēviņi ir homogamētiski. Ir arī citi dzimuma noteikšanas mehānismi. Tātad vairākiem kukaiņiem Y hromosomas nav. Šajā gadījumā viens no dzimumiem attīstās divu X hromosomu klātbūtnē, bet otrs - vienas X hromosomas klātbūtnē. Dažiem kukaiņiem dzimumu nosaka autosomu un dzimuma hromosomu skaita attiecība. Vairākos dzīvniekos t.s. dzimuma pārdefinēšana, kad atkarībā no vides faktoriem zigota kļūst vai nu par mātīti, vai par vīrieti. Dzimuma attīstībai augos ir tikpat dažādi ģenētiskie mehānismi kā dzīvniekiem.
Ar X hromosomu saistītās pazīmes. Ja gēns atrodas dzimuma hromosomā (to sauc par dzimumu saistītu), tad tā ekspresija pēcnācējos notiek saskaņā ar citiem noteikumiem, nevis autosomālajiem gēniem. Apsveriet gēnus, kas atrodas X hromosomā. Meita manto divas X hromosomas, vienu no mātes un vienu no tēva. Dēlam ir tikai viena X-hromosoma – no mātes; no tēva viņš saņem Y-hromosomu. Tāpēc tēvs savā X hromosomā esošos gēnus nodod tikai meitai, savukārt dēls tos nevar saņemt. Tā kā X hromosoma ir vairāk "bagātāka" ar gēniem, salīdzinot ar Y hromosomu, šajā ziņā meita ir ģenētiski līdzīgāka tēvam nekā dēlam; dēls vairāk līdzinās mātei, nevis tēvam.
Viena no vēsturiski vislabāk zināmajām ar dzimumu saistītajām pazīmēm cilvēkiem ir hemofilija, kas izraisa smagu asiņošanu no nelieliem iegriezumiem un plašus zilumus no sasitumiem. To izraisa recesīvi defektīva alēle 0, kas bloķē asins recēšanai nepieciešamā proteīna sintēzi. Šī proteīna gēns atrodas X hromosomā. Heterozigota sieviete +0 (+ nozīmē normālu aktīvo alēli, dominējoša attiecībā pret hemofilijas 0 alēli) ar hemofiliju nesaslimst, un arī viņas meitas, ja tēvam šīs patoloģijas nav. Tomēr viņas dēls var iegūt 0 alēli, un tad viņam attīstās hemofilija. X-hromosomu gēnu izraisītas recesīvās slimības sievietes skar daudz retāk nekā vīriešus, jo viņām šī slimība ir tikai tad, ja viņas ir homozigotas - recesīvās alēles klātbūtne katrā no divām homologajām X hromosomām; vīrieši saslimst ikreiz, kad viņu vienai X hromosomai ir bojāta alēle.
Saikne ar Y hromosomu.Informācija par gēniem, kas atrodas Y hromosomā, ir ļoti trūcīga. Tiek pieņemts, ka tas praktiski nesatur gēnus, kas nosaka šūnas funkcionēšanai nepieciešamo olbaltumvielu sintēzi. Bet tam ir galvenā loma vīriešu fenotipa attīstībā. Y hromosomas trūkums tikai vienas X hromosomas klātbūtnē noved pie t.s. Tērnera sindroms: sievietes fenotipa attīstība ar vāji attīstītām primārajām un sekundārajām seksuālajām īpašībām un citām novirzēm. Ir vīrieši ar papildu Y-hromosomu (XYY); tie ir gari, agresīvi un bieži vien ar neparastu uzvedību. Y hromosomā ir identificēti vairāki gēni, kas ir atbildīgi par specifisku enzīmu un hormonu sintēzes regulēšanu, un pārkāpumi tajos izraisa dzimuma attīstības patoloģijas. Pastāv vairākas morfoloģiskās pazīmes, kuras, domājams, nosaka Y hromosomas gēni; starp tiem ir ausu apmatojuma attīstība. Šādas pazīmes tiek pārraidītas tikai caur vīriešu līniju: no tēva uz dēlu.
ģenētiskā dzimuma noteikšana, nosaka dzimuma hromosomu kopums, atbalsta vienlīdzīgu mātīšu un tēviņu vairošanos. Patiešām, sieviešu olšūnas satur tikai X hromosomu, jo sievietēm ir XX genotips dzimuma hromosomās. Vīriešu genotips ir XY, un tāpēc meitenes vai zēna dzimšanu katrā gadījumā nosaka tas, vai spermā ir X vai Y hromosoma. Tā kā meiozes procesā hromosomām ir vienādas iespējas iekļūt gametā, tad puse no vīriešu dzimuma radītajām gametām satur X-hromosomu, bet puse - Y-hromosomu. Tāpēc paredzams, ka puse pēcnācēju būs viena dzimuma, bet puse – otra dzimuma.
Jāuzsver, ka iepriekš paredzēt zēna vai meitenes piedzimšanu nav iespējams, jo nav iespējams paredzēt, kura vīrieša dzimumšūna piedalīsies olšūnas apaugļošanā: tā, kura nesīs X vai Y hromosomu. . Tāpēc vairāk vai mazāk zēnu klātbūtne ģimenē ir nejaušības jautājums.
Saistītās mantojuma parādību analīze, šķērsošana, ģenētisko un citoloģisko karšu salīdzināšana ļauj formulēt galvenos hromosomu iedzimtības teorijas nosacījumus:
Gēni atrodas hromosomās. Turklāt dažādās hromosomas satur nevienlīdzīgu gēnu skaitu. Turklāt gēnu komplekts katrai nehomologajai hromosomai ir unikāls.
Alēlie gēni aizņem to pašu loci uz homologām hromosomām.
Gēni atrodas hromosomā lineārā secībā.
Vienas hromosomas gēni veido saiknes grupu, tas ir, tie tiek mantoti pārsvarā saistīti (kopīgi), kā rezultātā notiek dažu pazīmju saistīta pārmantošana. Saiknes grupu skaits ir vienāds ar konkrētās sugas hromosomu haploīdu skaitu (homogamētiskajā dzimumā) vai vairāk par 1 (heterogamētiskajā dzimumā).
Saikne tiek pārtraukta krustošanās rezultātā, kuras biežums ir tieši proporcionāls attālumam starp gēniem hromosomā (tādēļ saites stiprums ir apgriezti saistīts ar attālumu starp gēniem).
Katrai bioloģiskajai sugai ir raksturīgs noteikts hromosomu kopums - kariotips.
Saistīts mantojums
Neatkarīga funkciju kombinācija ( Mendeļa trešais likums) tiek veikta ar nosacījumu, ka gēni, kas nosaka šīs pazīmes, atrodas dažādos homologo hromosomu pāros. Tāpēc katrā organismā to gēnu skaitu, kas var neatkarīgi apvienoties meiozē, ierobežo hromosomu skaits. Tomēr organismā gēnu skaits ievērojami pārsniedz hromosomu skaitu. Piemēram, kukurūzā pirms molekulārās bioloģijas laikmeta tika pētīti vairāk nekā 500 gēnu, Drosophila mušai - vairāk nekā 1 tūkstotis, bet cilvēkiem - aptuveni 2 tūkstoši gēnu, savukārt tiem ir 10, 4 un 23 hromosomu pāri, attiecīgi. Tas, ka gēnu skaits augstākajos organismos ir vairāki tūkstoši, V. Setonam bija skaidrs jau 20. gadsimta sākumā. Tas ļāva pieņemt, ka katrā hromosomā ir lokalizēti daudzi gēni. Gēni, kas atrodas vienā hromosomā, veido saiknes grupu un tiek mantoti kopā.
Gēnu kopīga pārmantošana T. Morgan ierosināja nosaukt saistīts mantojums. Saiknes grupu skaits atbilst haploīdajam hromosomu skaitam, jo saiknes grupa sastāv no divām homologām hromosomām, kurās ir lokalizēti vieni un tie paši gēni. (Heterogamētiskā dzimuma indivīdiem, piemēram, zīdītāju tēviņiem, faktiski ir vēl viena saiknes grupa, jo X un Y hromosomas satur dažādus gēnus un pārstāv divas dažādas saiknes grupas. Tādējādi sievietēm ir 23 saišu grupas, bet vīriešiem - 24).
Saistīto gēnu pārmantošanas veids atšķiras no gēnu mantojuma, kas atrodas dažādos homologo hromosomu pāros. Tātad, ja ar neatkarīgu kombināciju diheterozigots indivīds veido četru veidu gametas (AB, Ab, aB un ab) vienādos daudzumos, tad ar saistītu iedzimtību (ja nav šķērsošanas), viens un tas pats diheterozigots veido tikai divu veidu gametas. gametas: (AB un ab) arī vienādos daudzumos. Pēdējie atkārto gēnu kombināciju vecāku hromosomā.
Tomēr tika konstatēts, ka līdztekus parastajām (nekrustojošām) gametām ar jaunām gēnu kombinācijām - Ab un aB, kas atšķiras no vecāku hromosomu gēnu kombinācijām, rodas arī citas (crossover) gametas. Šādu gametu veidošanās iemesls ir homologu hromosomu sekciju apmaiņa vai šķērsojot.
Pāreja notiek mejozes I fāzē laikā konjugācijas homologās hromosomas. Šajā laikā divu hromosomu daļas var krustoties un apmainīties ar savām daļām. Tā rezultātā rodas kvalitatīvi jaunas hromosomas, kas satur gan mātes, gan tēva hromosomu sekcijas (gēnus). Indivīdus, kas iegūti no šādām gametām ar jaunu alēļu kombināciju, sauc par krustošanos vai rekombinantiem.
Divu vienā un tajā pašā hromosomā esošo gēnu krustošanās biežums (procentos) ir proporcionāls attālumam starp tiem. Divu gēnu krustošanās notiek retāk, jo tuvāk tie atrodas viens otram. Palielinoties attālumam starp gēniem, arvien vairāk palielinās iespējamība, ka šķērsošana tos atdalīs divās dažādās homologās hromosomās.
Attālums starp gēniem raksturo to saiknes stiprumu. Ir gēni ar lielu saiknes procentuālo daļu un tādi, kuros saikne gandrīz netiek atklāta. Tomēr ar saistītu mantojumu maksimālā krustošanas frekvence nepārsniedz 50%. Ja tas ir augstāks, tad starp alēļu pāriem ir brīva kombinācija, kas nav atšķirama no neatkarīgas mantojuma.
Šķērsošanas bioloģiskā nozīme ir ārkārtīgi augsta, jo ģenētiskā rekombinācijaļauj izveidot jaunas, iepriekš neeksistējošas gēnu kombinācijas un tādējādi palielināt iedzimtību mainīgums, kas sniedz plašas iespējas organisma adaptācijai dažādos vides apstākļos. Cilvēks speciāli diriģē hibridizācija lai iegūtu nepieciešamās kombinācijas lietošanai audzēšanas darbs.
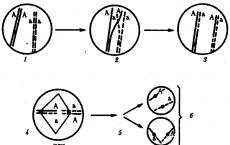
Šķērsojot.Šis process notiek mejozes I fāzē laikā, kad konjugācijas rezultātā homologās hromosomas ir cieši savienotas un veido bivalentus. Šķērsošanas laikā notiek atbilstošo sekcijas apmaiņa starp savstarpēji savijas homologu hromosomu hromatīdiem (3.72. att.). Šis process nodrošina gēnu tēva un mātes alēļu rekombināciju katrā saišu grupā. Dažādos gametu prekursoros šķērsošana notiek dažādās hromosomu daļās, kā rezultātā hromosomās rodas dažādas vecāku alēļu kombinācijas.
Rīsi. 3.72. Šķērsošana kā gametu ģenētiskās daudzveidības avots:
I - vecāku gametu apaugļošana a un b c zigotu veidošanās iekšā;II - gametoģenēze organismā, kas attīstījies no zigotas iekšā;G- pāreja starp homologiem profāzē es; d - šūnas, kas veidojas pēc 1. meiotiskā dalīšanās; e, w - šūnas, kas veidojas pēc mejozes 2. dalīšanās ( e - nešķērsojošas gametas ar sākotnējām vecāku hromosomām; labi - krusteniskās gametas ar iedzimta materiāla rekombināciju homologās hromosomās)
Ir skaidrs, ka šķērsošana kā rekombinācijas mehānisms ir efektīvs tikai tad, ja atbilstošos tēva un mātes hromosomu gēnus attēlo dažādas alēles. Absolūti identiskas saišu grupas krustošanās laikā nedod jaunas alēļu kombinācijas.
Mejozes laikā krustošanās notiek ne tikai dzimumšūnu prekursoros. To novēro arī somatiskajās šūnās mitozes laikā. Somatiskā krustošanās ir aprakstīta Drosophila un dažās pelējuma sugās. To veic mitozes laikā starp homologām hromosomām, bet tā biežums ir 10 000 reižu mazāks nekā meiotiskās šķērsošanas biežums, no kura mehānisma tas neatšķiras. Mitotiskās šķērsošanas rezultātā parādās somatisko šūnu kloni, kas atšķiras pēc atsevišķu gēnu alēļu satura tajos. Ja zigotas genotipā šis gēns ir attēlots ar divām dažādām alēlēm, tad somatiskās krustošanas rezultātā var parādīties šūnas ar vienādām šī gēna tēva vai mātes alēlēm (3.73. att.).

Rīsi. 3.73. Šķērsošana somatiskajās šūnās:
1 - somatiskā šūna, kuras homologajās hromosomās gēnu A pārstāv divas dažādas alēles (A un a); 2 - šķērsošana; 3 - atbilstošo sekciju apmaiņas rezultāts starp homologām hromosomām; 4 - homologu izvietojums skaldīšanas vārpstas ekvatoriālajā plaknē mitozes metafāzē (divi varianti); 5 - meitas šūnu veidošanās; 6 - gēnam A heterozigotu šūnu veidošanās, kas alēļu kopuma (Aa) ziņā ir līdzīgas mātes šūnai; 7 - gēnam A homozigotu šūnu veidošanās, kas atšķiras no mātes šūnas alēļu komplektā (AA vai aa)
19. gadsimta vidū, kad G. Mendelis veica savus eksperimentus un formulēja universālas un fundamentālas nozīmes modeļus ģenētikas un bioloģijas attīstībai kopumā, zinātniskās zināšanas vēl nebija pietiekamas, lai izprastu mantojuma mehānismus. Tāpēc Mendeļa darbs daudzus gadus nebija pieprasīts. Tomēr līdz 20. gadsimta sākumam. situācija bioloģijā ir radikāli mainījusies.
Tika atklāta mitoze un mejoze, no jauna tika atklāti Mendeļa likumi. Neatkarīgi viens no otra pētnieki Vācijā un ASV ierosināja, ka iedzimtie faktori atrodas hromosomās. 1906. gadā R. Penets pirmo reizi aprakstīja Mendeļa likuma par divu pazīmju neatkarīgu pārmantošanu pārkāpumu. Iestudējot klasisko dihibrīdo saldo zirņu augu krustojumu, kas atšķiras pēc ziedu krāsas un ziedputekšņu formas, Penets otrajā paaudzē nesaņēma gaidīto šķelšanos 9:3:3:1. F2 hibrīdiem bija tikai vecāku fenotipi proporcijā 3:1, t.i., pazīmju pārdale nenotika.
Pamazām arvien vairāk sakrājās tādi izņēmumi, kas nepakļāvās patstāvīgās mantošanas likumam. Radās jautājums, kā tieši gēni atrodas hromosomās? Galu galā pazīmju skaits un līdz ar to arī gēnu skaits katrā organismā ir daudz lielāks nekā hromosomu skaits. Tas nozīmē, ka katrā hromosomā ir daudz gēnu, kas atbild par dažādām pazīmēm. Kā tiek mantoti gēni, kas atrodas vienā hromosomā?
■ T. Morgana darbs. Uz šiem jautājumiem atbildēja amerikāņu zinātnieku grupa Tomasa Hanta Morgana (1866-1945) vadībā. Strādājot pie ļoti ērta ģenētiska objekta – augļmušas Drosophila, viņi lieliski paveica gēnu pārmantojamības izpēti.
Zinātnieki to ir atklājuši gēni, kas atrodas vienā hromosomā, tiek mantoti kopā, t.i., saistīti.Šī parādība ir nosaukta Morgan likums, vai saistītās mantošanas likums . Tiek sauktas gēnu grupas, kas atrodas vienā hromosomā sajūga grupa. Tā kā homologās hromosomās atrodas identiski gēni, saišu grupu skaits ir vienāds ar hromosomu pāru skaitu, t.i., hromosomu haploīdo skaitu. Cilvēkam ir 23 pāri hromosomu un līdz ar to 23 saišu grupas, sunim ir 39 pāri hromosomu un 39 saišu grupas, zirnī ir 7 pāri hromosomu un 7 saišu grupas utt. Jāpiebilst, ka Mendelam bija pārsteidzoši paveicies veidojot dihibrīdos krustojumus: gēni, kas atbild par dažādām pazīmēm (zirņu krāsa un forma), atradās dažādās hromosomās. Varēja būt citādi, un tad neatkarīgas šķelšanās modelis nebūtu atklāts.
T. Morgana grupas darba rezultāts bija 1911. gada izveide hromosomu iedzimtības teorija.
Apsveriet mūsdienu hromosomu iedzimtības teorijas galvenos noteikumus.
Iedzimtības vienība ir gēns, kas ir hromosomas segments.
Gēni atrodas hromosomās stingri noteiktās vietās (lokusos), un alēlie gēni (atbildīgi par vienas pazīmes attīstību) atrodas tajos pašos homologo hromosomu lokusos.
Gēni atrodas hromosomās lineārā secībā, tas ir, viens pēc otra.
■Sajūga kļūme. Tomēr dažos krustojumos, analizējot tajā pašā hromosomā esošo gēnu pārmantošanu, tika konstatēts saiknes traucējums. Izrādījās, ka dažkārt pārī savienotas homologas hromosomas var savstarpēji apmainīties ar identiskiem homologiem reģioniem. Lai tas notiktu, hromosomām jāatrodas tiešā tuvumā. Šo homologo hromosomu pagaidu pāru pieeju sauc par konjugāciju. Šajā gadījumā hromosomas var apmainīties ar lokusiem, kas atrodas viens pret otru un satur vienus un tos pašus gēnus. Šo parādību sauc par šķērsošanu.
Atgādiniet meiozes dalīšanos, kuras laikā veidojas dzimumšūnas. Pirmā meiotiskā dalījuma profāzē, veidojoties bivalentam (tetradam), dubultotām homologām hromosomām stāvot paralēli viena otrai, var notikt līdzīga apmaiņa. Šāds notikums noved pie ģenētiskā materiāla rekombinācijas, palielina pēcnācēju daudzveidību, t.i., palielina iedzimto mainīgumu un tāpēc spēlē nozīmīgu lomu evolūcijā.
■ Ģenētiskās kartes. Alēļu gēnu apmaiņas fenomens starp homologām hromosomām palīdzēja zinātniekiem noteikt katra gēna atrašanās vietu hromosomā, tas ir, izveidot ģenētiskās kartes. Hromosomas ģenētiskā karte ir diagramma par gēnu savstarpējo izvietojumu, kas atrodas vienā hromosomā, t.i., vienā saišu grupā. Par šādu karšu uzbūvi ir liela interese gan fundamentāliem pētījumiem, gan dažādu praktisku problēmu risināšanai. Piemēram, cilvēka hromosomu ģenētiskās kartes ir ļoti svarīgas, lai diagnosticētu vairākas smagas iedzimtas slimības.
Šobrīd vienkāršas ģenētiskās kartes tiek aizstātas ar molekulārām ģenētiskajām kartēm, kurās ir informācija par gēnu nukleotīdu sekvencēm.
Jautājumi paškontrolei
1. Kas ir saistītais mantojums?
2. Kas ir gēnu saišu grupas?
3. Kāds ir gēnu saites traucējumu cēlonis?
4. Kāda ir alēlo gēnu apmaiņas bioloģiskā nozīme starp homologām hromosomām?
5. Vai saistītā mantojuma teorija ir apstiprināta citoloģiski?
Pēc G. Mendela domām, gēni tiek mantoti neatkarīgi un atrodas dažādās hromosomās. Diheterozigotu AaBb citoloģiskās formulas veidā var attēlot šādi:
Mejozes laikā tas veido 4 veidu gametas: AB, Ab, aB, ab.
1911.–1912 Amerikāņu ģenētiķis T. Morgans pierādīja, ka gēni, kas atrodas vienā hromosomā, var tikt mantoti saistīti (kopā). Tas ir saistīts ar faktu, ka ir daudz vairāk gēnu nekā hromosomu. Gēnu līdzmantošana ierobežojot to brīvo kombināciju, piesauca T. Morgans gēnu saikne vai saistīta mantojums. Gadījumā, ja gēni ir iedzimti saistīti, AaBb diheterozigota genotipu, izmantojot citoloģisko formulu, var attēlot šādi:
Meiozes laikā, ja ir pilnīga gēnu saite, veidojas 2 veidu gametas - AB, ab. Šādas gametas sauc par nekrosoveriem.
T. Morgana un viņa studentu pētījumi parādīja, ka homologā hromosomu pārī regulāri notiek gēnu apmaiņa. Procesu, kurā notiek homologu hromosomu identisku sekciju apmaiņa ar tajās esošajiem gēniem, sauc par hromosomu dekusāciju, vai šķērsojot.Šķērsošana nodrošina jaunas gēnu kombinācijas, kas atrodas homologās hromosomās . Tāpēc krustošanās fenomena dēļ papildus nekrosoveram ir arī krustojošās gametas - Ab, aB.
T. Morgana likums ir formulēts šādi: gēni, kas atrodas vienā hromosomā, ir iedzimti saistīti (kopā), un saites stiprums ir atkarīgs no attāluma starp gēniem.
T. Morgana hromosomu teorijas galvenie nosacījumi:
1) gēni ir lokalizēti hromosomās, un dažādas hromosomas satur nevienlīdzīgu gēnu skaitu. Gēnu kopums katrā no nehomologajām hromosomām ir unikāls;
2) gēni atrodas hromosomā noteiktā lineārā secībā;
3) gēni veido saiknes grupas un var tikt mantoti saistīti. Saiknes grupu skaits atbilst haploīdajam hromosomu kopumam (cilvēkam ir 23 saišu grupas);
4) starp homologām hromosomām iespējama krustošanās (crossover) un alēlo gēnu apmaiņa, un krustojums notiek, kad starp gēniem ir attālums;
5) attālums starp gēniem hromosomā ir proporcionāls krustošanās biežumam un ir izteikts krustošanās procentos.
Gēnu krustošanās procentuālo daļu aprēķina pēc formulas:
X= ––––
a ir vienas klases krosoveru indivīdu skaits; b ir citas klases krosoveru indivīdu skaits;
n ir īpatņu kopējais skaits; Х – % no krustošanās starp gēniem (attālums starp gēniem % no krustojuma).
Attālums starp gēniem tiek mērīts īpašās vienībās - morganīdi. 1 morganīds (1 M) = 1% šķērsošana.
Ģenētiskajos pētījumos tika izmantota augļu muša, kuras kariotips sastāv no 8 hromosomām, auglība un daudzas alternatīvas pazīmes. T. Morgans apsvēra situāciju, kad pilnīga gēnu saistība un gēnu sasaistes pārkāpums (šķērsošana) rodas indivīdā vienlaikus. Pirmajā posmā krustošanai tika ņemti homozigoti indivīdi (P ♀AABB x ♂aabb). Tā rezultātā pirmajā paaudzē visi indivīdi bija diheterozigoti (AaBb).
Gēnu iezīme
Pelēka ķermeņa krāsa
un tumša ķermeņa krāsa
Garos spārnos
b rudimentāri spārni.
Pilna gēnu saistība
P ♀ ══x ♂ ══
G AB, ab ab - vīriešu dzimumšūnas
(Sieviešu dzimumšūnas, kas nav krustojušās)
F 1 ══ ; ══
Ar pilnīgu gēnu saikni tiek novērota 1:1 sadalīšanās. Kopumā tika iegūti 83% indivīdu, kuri mantojuši savu vecāku fenotipu, jo tie veidojās no ne-crossover gametām. No tiem indivīdi ar pelēku ķermeni, normāliem spārniem veidoja 41,5% un indivīdi ar melnu ķermeni, rudimentāriem spārniem arī veidoja 41,5%.
Gēnu saiknes traucējumi
P ♀ ══ x ♂ ══
G Ab, aB ab - vīriešu dzimumšūnas
(Sievišķās dzimumšūnas)
F 1 ══; ══
Ar nepilnīgu gēnu saiti tiek novērota arī 1:1 sadalīšanās. Kopumā tika iegūti 17% indivīdu ar rekombinantiem fenotipiem, jo tie veidojās no krustojošām gametām. Indivīdi ar jaunu pazīmju kombināciju tiek saukti rekombinanti. Starp rekombinantiem mušas ar pelēku ķermeni, rudimentāri spārni veidoja 8,5%, bet ar melnu ķermeni arī normāli spārni veidoja 8,5%.
1. jautājums. Kas ir saistīta mantošana?
Saistītā mantošana ir vienā un tajā pašā hromosomā (t.i., vienā DNS molekulā) esošo gēnu kopīgs pārmantojums. Piemēram, saldajos zirņos šādi izkārtojušies gēni, kas nosaka ziedu krāsu un ziedputekšņu formu. Tie tiek mantoti sasaistītā veidā, tāpēc, krustojot otrās paaudzes hibrīdus, vecāku fenotipi veidojas attiecībā 3:1, un sadalīšanās 9:3:3:1, kas raksturīga dihibrīda krustojumam ar neatkarīgu pārmantošanu. neparādās.
2. jautājums. Kas ir gēnu saišu grupas?
Saiknes grupa ir gēnu kolekcija, kas atrodas vienā hromosomā. Tā kā homologās hromosomas satur vienādus gēnus, saišu grupu skaits ir vienāds ar haploīdo hromosomu skaitu (23 cilvēkiem, 7 zirņiem, 4 Drosophila).
3. jautājums. Kāds ir gēnu saites traucējumu cēlonis?
Gēnu saiknes pārtraukšanas iemesls ir homologu hromosomu sekciju apmaiņa meiotiskās dalīšanas I fāzē. Atgādiniet, ka šajā posmā pārī savienotas hromosomas ir konjugētas, veidojot tā sauktās divvērtīgās joslas. Bivalentu veidošanās var izraisīt hromosomu krustojumu, kas rada iespēju apmainīties ar homologiem DNS reģioniem. Ja tā notiek, tad saišu grupas maina savu saturu (tās satur citas to pašu gēnu alēles) un pēcnācējos var parādīties indivīdi ar fenotipu, kas atšķiras no vecākiem.
4. jautājums. Kāda ir alēlo gēnu apmaiņas bioloģiskā nozīme starp homologām hromosomām?materiāls no vietnes
Hromosomu krustošanās (citādi - krustošanās) noved pie ģenētiskā materiāla rekombinācijas un jaunu gēnu alēļu kombināciju veidošanās no saiknes grupas. Tajā pašā laikā palielinās pēcnācēju daudzveidība, t.i., palielinās iedzimta mainība, kam ir liela evolucionāra nozīme. Patiešām, ja, piemēram, Drosofilā gēni, kas nosaka ķermeņa krāsu un spārnu garumu, atrodas vienā hromosomā, tad, krustojot tīras pelēko mušu līnijas ar normāliem spārniem un melnajām mušām ar saīsinātiem spārniem, ja nav šķērsošanas, mēs nekad neiegūsim citus fenotipus. Hromosomu šķērsošanas esamība ļauj parādīties (dažos procentos gadījumu) pelēkām mušām ar īsiem spārniem un melnām mušām ar normāliem spārniem.
5. jautājums. Vai saistītā mantojuma teorija ir apstiprināta citoloģiski?
Tomasa Hanta Morgana (1866-1945) teoriju par saistīto pārmantošanu apstiprina citoloģiskie novērojumi. Tika parādīts, ka dalīšanās laikā hromosomas pilnībā novirzās uz dažādiem šūnas poliem. Līdz ar to gēni, kas meiozes laikā atrodas vienā un tajā pašā hromosomā, ietilpst vienā gametā, tas ir, tie patiešām tiek mantoti saistītā veidā.
Vai neatradāt to, ko meklējāt? Izmantojiet meklēšanu