Teoria moștenirii legate este confirmată citologic? Moștenirea legată a genelor. Teoria cromozomală a eredităţii T. Morgan. Revizuiți întrebările și temele
Moștenirea legată - moştenirea trăsăturilor ale căror gene sunt localizate pe acelaşi cromozom. Puterea legăturii dintre gene depinde de distanța dintre ele: cu cât genele sunt mai departe una de cealaltă, cu atât este mai mare frecvența de încrucișare și invers. Alături de trăsăturile care sunt moștenite independent, trebuie să existe și cele care sunt moștenite legate între ele, deoarece sunt determinate de gene situate pe același cromozom. Se formează astfel de gene grup de ambreiaj. Numărul de grupuri de legătură în organismele unei anumite specii este egal cu numărul de cromozomi din setul haploid (de exemplu, la Drosophila 1 pereche = 4, la om 1 pereche = 23).
Prindere totală- un fel de moștenire legată, în care genele trăsăturilor analizate sunt situate atât de aproape una de cealaltă încât trecerea între ele devine imposibilă.
Ambreiaj incomplet- un tip de moștenire legată, în care genele trăsăturilor analizate sunt situate la o anumită distanță unele de altele, ceea ce face posibilă încrucișarea între ele.
(Încrucișarea gameților- gameți, în procesul de formare a căror încrucișare a avut loc. De regulă, gameții încrucișați reprezintă o mică parte din numărul total de gameți.
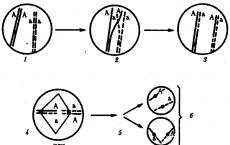
Trecere peste- schimb de secțiuni de cromozomi omologi în procesul de diviziune celulară, în principal în profaza primei diviziuni meiotice, uneori în mitoză. Experimentele lui T. Morgan, K. Bridges și A. Sturtevant au arătat că nu există o legătură absolut completă a genelor, în care genele ar fi întotdeauna transmise împreună. Probabilitatea ca două gene situate pe același cromozom să nu diverge în timpul meiozei variază de la 1 la 0,5. În natură, legătura incompletă predomină datorită încrucișării cromozomilor omologi și recombinării genelor. Modelul citologic de încrucișare a fost descris pentru prima dată de omul de știință danez F. Janssens.
Încrucișarea are loc numai atunci când genele sunt într-o stare heterozigotă (AB / av). Dacă genele sunt într-o stare homozigotă (AB/AB sau aB/aB), schimbul de regiuni identice nu dă noi combinații de gene în gameți și într-o generație. Frecvența (procentul) de încrucișare între gene depinde de distanța dintre ele: cu cât sunt mai îndepărtate una de cealaltă, cu atât se produce mai des încrucișarea. T. Morgan a propus să măsoare distanța dintre gene prin încrucișare ca procent, conform formulei:
N1/N2 X 100 = % Crossover,
unde N1 este numărul total de indivizi din F;
N2 este numărul total de indivizi încrucișați.
Segmentul cromozomului, pe care are loc 1% de încrucișare, este egal cu o morganidă (o măsură condiționată a distanței dintre gene). Frecvența de încrucișare este utilizată pentru a determina poziția relativă a genelor și distanța dintre ele. Pentru a construi o hartă genetică umană, se folosesc tehnologii noi, în plus, s-au construit hărți citogenetice ale cromozomilor.
Există mai multe tipuri de traversări: duble, multiple (complexe), neregulate, inegale.
Încrucișarea duce la o nouă combinație de gene, provocând o schimbare a fenotipului. În plus, împreună cu mutațiile, este un factor important în evoluția organismelor.)
Rezultatul cercetărilor lui T. Morgan a fost creația sa a teoriei cromozomiale a eredității:
· genele sunt localizate pe cromozomi; diferiți cromozomi conțin un număr inegal de gene; setul de gene pentru fiecare dintre cromozomii neomologi este unic;
· fiecare genă are o locație specifică (locus) pe cromozom; genele alelice sunt localizate în loci identici ai cromozomilor omologi;
· genele sunt localizate pe cromozomi într-o anumită secvență liniară;
· genele situate pe același cromozom sunt moștenite împreună, formând un grup de legătură; numărul de grupuri de legătură este egal cu setul haploid de cromozomi și este constant pentru fiecare tip de organism;
· legarea genelor poate fi ruptă în timpul procesului de încrucișare, ceea ce duce la formarea de cromozomi recombinanți; frecvența încrucișării depinde de distanța dintre gene: cu cât distanța este mai mare, cu atât valoarea încrucișării este mai mare;
· fiecare specie are un set de cromozomi caracteristici doar pentru ea - un cariotip.
Moștenirea sexului și a trăsăturilor legate de sex. Cromozomii sexuali și rolul lor în determinarea sexului. Moștenirea sexuală. Sexul unui individ este o trăsătură complexă, formată atât prin acțiunea genelor, cât și prin condițiile dezvoltării. La om, una dintre cele 23 de perechi de cromozomi este cromozomii sexuali, notați X și Y. Femeile sunt sex homogametic, adică. Au doi cromozomi X, unul de la mama și unul de la tatăl lor. Masculii sunt heterogametici, au un cromozom Y X-un, iar X se transmite de la mamă, iar Y de la tată. Rețineți că sexul heterogametic nu este întotdeauna neapărat masculin; de exemplu, la păsări, acestea sunt femele, în timp ce masculii sunt homogametici. Există și alte mecanisme de determinare a sexului. Deci, la o serie de insecte, cromozomul Y este absent. În acest caz, unul dintre sexe se dezvoltă în prezența a doi cromozomi X, iar celălalt - în prezența unui cromozom X. La unele insecte, sexul este determinat de raportul dintre numărul de autozomi și cromozomi sexuali. Într-un număr de animale, așa-numitele. redefinirea sexului, când, în funcție de factorii de mediu, zigotul se dezvoltă fie într-o femeie, fie într-un mascul. Dezvoltarea sexului la plante are mecanisme genetice la fel de diverse ca și la animale.
Trăsături legate de cromozomul X. Dacă gena este localizată pe cromozomul sexual (se numește sex-linked), atunci exprimarea ei în descendenți urmează alte reguli decât pentru genele autozomale. Luați în considerare genele situate pe cromozomul X. O fiică moștenește doi cromozomi X, unul de la mama și unul de la tatăl ei. Fiul are un singur cromozom X - de la mamă; de la tatăl său primește un cromozom Y. Prin urmare, tatăl transmite genele de pe cromozomul său X doar fiicei sale, în timp ce fiul nu le poate primi. Deoarece cromozomul X este mai „bogat” în gene în comparație cu cromozomul Y, în acest sens, fiica se aseamănă genetic mai mult cu tatăl decât cu fiul; fiul seamănă mai mult cu mama lui decât cu tatăl său.
Una dintre cele mai cunoscute trăsături istorice legate de sex la oameni este hemofilia, care are ca rezultat sângerare severă de la tăieturi minore și vânătăi extinse de la vânătăi. Este cauzată de o alelă recesivă defectuoasă 0, care blochează sinteza unei proteine necesare coagulării sângelui. Gena acestei proteine este localizată pe cromozomul X. O femeie heterozigotă +0 (+ înseamnă alela activă normală, dominantă în raport cu alela hemofilie 0) nu face hemofilie, iar fiicele ei, de asemenea, dacă tatăl nu are această patologie. Cu toate acestea, fiul ei poate obține alela 0 și apoi dezvoltă hemofilie. Bolile recesive cauzate de genele cromozomului X afectează femeile mult mai rar decât bărbații, întrucât au boala doar atunci când sunt homozigoți – prezența unei alele recesive în fiecare dintre cei doi cromozomi X omologi; bărbații se îmbolnăvesc ori de câte ori unicul lor cromozom X poartă alela defectuoasă.
Legătura cu cromozomul Y.Informațiile despre genele situate pe cromozomul Y sunt foarte rare. Se presupune că practic nu poartă genele care determină sinteza proteinelor necesare funcționării celulei. Dar joacă un rol cheie în dezvoltarea fenotipului masculin. Absența unui cromozom Y în prezența unui singur cromozom X duce la așa-numitul. Sindromul Turner: dezvoltarea unui fenotip feminin cu caracteristici sexuale primare și secundare slab dezvoltate și alte anomalii. Există bărbați cu un cromozom Y suplimentar (XYY); sunt înalți, agresivi și au adesea un comportament anormal. În cromozomul Y, au fost identificate mai multe gene care sunt responsabile de reglarea sintezei unor enzime și hormoni specifici, iar încălcările acestora duc la patologii ale dezvoltării sexuale. Există o serie de trăsături morfologice despre care se crede că sunt determinate de genele de pe cromozomul Y; printre acestea se numără și dezvoltarea părului urechii. Astfel de semne se transmit doar prin linia masculină: de la tată la fiu.
determinarea genetică a sexului, determinat de setul de cromozomi sexuali, susține reproducerea egală a femelelor și masculilor. Într-adevăr, ouăle feminine conțin doar cromozomul X, deoarece femeile au genotipul XX pe cromozomii sexuali. Genotipul bărbaților este XY și, prin urmare, nașterea unei fete sau a unui băiat în fiecare caz este determinată de dacă spermatozoidul poartă cromozomul X sau Y. Deoarece în procesul de meioză cromozomii au șanse egale de a intra în gamet, atunci jumătate din gameții produși de indivizii de sex masculin conțin cromozomul X și jumătate din cromozomul Y. Prin urmare, jumătate din urmași sunt de așteptat să fie de un sex și jumătate de celălalt.
Trebuie subliniat faptul că este imposibil să se prezică în avans nașterea unui băiat sau a unei fete, deoarece este imposibil să se prezică care celulă germinală masculină va participa la fertilizarea ovulului: cea care poartă cromozomul X sau Y. . Prin urmare, prezența mai multor sau mai puțini băieți în familie este o chestiune de întâmplare.
Analiza fenomenelor de moștenire legată, crossing over, compararea hărților genetice și citologice ne permit să formulăm principalele prevederi ale teoriei cromozomiale a eredității:
Genele sunt localizate pe cromozomi. Mai mult, diferiți cromozomi conțin un număr inegal de gene. În plus, setul de gene pentru fiecare dintre cromozomii neomologi este unic.
Genele alelice ocupă la fel loci pe cromozomi omologi.
Genele sunt localizate pe cromozom într-o secvență liniară.
Genele unui cromozom formează un grup de legătură, adică sunt moștenite predominant legate (în comun), datorită cărora are loc moștenirea legată a unor trăsături. Numărul de grupuri de legături este egal cu numărul haploid de cromozomi ale unei specii date (la sexul homogametic) sau mai mult cu 1 (la sexul heterogametic).
Legătura este întreruptă ca urmare a încrucișării, a cărei frecvență este direct proporțională cu distanța dintre genele din cromozom (prin urmare, puterea legăturii este invers proporțională cu distanța dintre gene).
Fiecare specie biologică este caracterizată de un anumit set de cromozomi - cariotip.
Moștenirea legată
Combinație independentă de caracteristici ( A treia lege a lui Mendel) se efectuează cu condiția ca genele care determină aceste trăsături să fie în diferite perechi de cromozomi omologi. Prin urmare, în fiecare organism, numărul de gene care se pot combina independent în meioză este limitat de numărul de cromozomi. Cu toate acestea, într-un organism, numărul de gene depășește semnificativ numărul de cromozomi. De exemplu, în porumb înainte de era biologiei moleculare, au fost studiate peste 500 de gene, la musca Drosophila - mai mult de 1 mie, iar la oameni - aproximativ 2 mii de gene, în timp ce au 10, 4 și 23 de perechi de cromozomi, respectiv. Faptul că numărul de gene în organismele superioare este de câteva mii era deja clar pentru W. Setton la începutul secolului al XX-lea. Acest lucru a dat motive să presupunem că multe gene sunt localizate în fiecare cromozom. Genele situate pe același cromozom formează un grup de legătură și sunt moștenite împreună.
Moștenirea comună a genelor T. Morgan a propus să numească moștenire legată. Numărul de grupuri de legătură corespunde numărului haploid de cromozomi, deoarece grupul de legătură este format din doi cromozomi omologi în care sunt localizate aceleași gene. (La indivizii de sex heterogametic, de exemplu, mamiferele masculi, există de fapt încă un grup de legătură, deoarece cromozomii X și Y conțin gene diferite și reprezintă două grupuri de legătură diferite. Astfel, femeile au 23 de grupuri de legătură, iar la bărbați - 24).
Modul de moștenire a genelor legate diferă de moștenirea genelor situate în diferite perechi de cromozomi omologi. Deci, dacă, cu o combinație independentă, un individ diheterozigot formează patru tipuri de gameți (AB, Ab, aB și ab) în cantități egale, atunci cu moștenire legată (în absența încrucișării), același diheterozigot formează doar două tipuri de gameți: (AB și ab) tot în cantități egale. Acestea din urmă repetă combinația de gene din cromozomul părintelui.
S-a constatat însă că, pe lângă gameții obișnuiți (neîncrucișați), apar și alți gameți (încrucișați) cu noi combinații de gene - Ab și aB, care diferă de combinațiile de gene din cromozomii părintelui. Motivul formării unor astfel de gameți este schimbul de secțiuni de cromozomi omologi sau trecere peste.
Încrucișarea are loc în profaza I a meiozei în timpul conjugări cromozomi omologi. În acest moment, părți din doi cromozomi se pot trece și schimba părțile lor. Ca rezultat, apar cromozomi noi din punct de vedere calitativ, care conțin secțiuni (gene) atât ale cromozomilor materni, cât și ale celor paterni. Indivizii care sunt obținuți din astfel de gameți cu o nouă combinație de alele se numesc crossing-over sau recombinanți.
Frecvența (procentul) de încrucișare între două gene situate pe același cromozom este proporțională cu distanța dintre ele. Încrucișarea între două gene are loc mai puțin frecvent cu cât acestea sunt mai aproape una de cealaltă. Pe măsură ce distanța dintre gene crește, probabilitatea ca încrucișarea le va separa pe doi cromozomi omologi diferiți crește din ce în ce mai mult.
Distanța dintre gene caracterizează puterea legăturii lor. Există gene cu un procent mare de legare și acelea în care legarea aproape nu este detectată. Cu toate acestea, cu moștenirea legată, frecvența maximă de încrucișare nu depășește 50%. Dacă este mai mare, atunci există o combinație liberă între perechi de alele, care nu se distinge de moștenirea independentă.
Semnificația biologică a crossing over este extrem de mare, din moment ce genetică recombinare vă permite să creați combinații noi, inexistente anterior, de gene și, prin urmare, să creșteți ereditar variabilitate, care oferă oportunități ample de adaptare a organismului în diferite condiții de mediu. O persoană conduce în mod special hibridizareîn vederea obţinerii combinaţiilor necesare utilizării în munca de reproducere.
Trecere peste. Acest proces are loc în profaza I a meiozei într-un moment în care cromozomii omologi sunt strâns reuniți ca rezultat al conjugării și formează bivalenți. În timpul încrucișării, secțiunile corespunzătoare sunt schimbate între cromatidele cromozomilor omologi care se întrepătrund reciproc (Fig. 3.72). Acest proces asigură recombinarea alelelor paterne și materne ale genelor în fiecare grup de legătură. La diferiți precursori ai gameților, încrucișarea are loc în diferite părți ale cromozomilor, rezultând o mare varietate de combinații de alele parentale în cromozomi.
Orez. 3.72. Încrucișarea ca sursă a diversității genetice a gameților:
I - fertilizarea gametilor parentali a și b c formarea zigotului în;II - gametogeneza într-un organism dezvoltat dintr-un zigot în;G- trecerea între omologi în profază eu; d - celulele formate după prima diviziune meiotică; e, w - celulele formate după a doua diviziune a meiozei ( e - gameți neîncrucișați cu cromozomi parentali originali; bine - gameți încrucișați cu recombinarea materialului ereditar în cromozomi omologi)
Este clar că încrucișarea ca mecanism de recombinare este eficientă numai atunci când genele corespunzătoare ale cromozomilor patern și materni sunt reprezentate de alele diferite. Grupurile de legătură absolut identice în timpul încrucișării nu dau noi combinații de alele.
Încrucișarea are loc nu numai în precursorii celulelor germinale în timpul meiozei. Se observă și în celulele somatice în timpul mitozei. Încrucișarea somatică a fost descrisă la Drosophila și la unele specii de mucegai. Se efectuează în timpul mitozei între cromozomi omologi, dar frecvența sa este de 10.000 de ori mai mică decât frecvența încrucișării meiotice, de al cărui mecanism nu diferă. Ca urmare a încrucișării mitotice, apar clone de celule somatice care diferă în conținutul alelelor individuale ale genelor din ele. Dacă în genotipul zigotului această genă este reprezentată de două alele diferite, atunci ca urmare a încrucișării somatice pot apărea celule cu aceleași alele fie paterne, fie materne ale acestei gene (Fig. 3.73).

Orez. 3.73 Încrucișarea în celulele somatice:
1 - o celula somatica, in cromozomii omologi ai carei gena A este reprezentata de doua alele diferite (A si a); 2 - trecere peste; 3 - rezultatul schimbului de secțiuni corespunzătoare între cromozomi omologi; 4 - localizarea omologilor în planul ecuatorial al fusului de fisiune în metafaza mitozei (două variante); 5 - formarea celulelor fiice; 6 - formarea de celule heterozigote pentru gena A, similare cu celula mamă în ceea ce privește un set de alele (Aa); 7 - formarea celulelor homozigote pentru gena A, care diferă de celula mamă în setul de alele (AA sau aa)
La mijlocul secolului al XIX-lea, când G. Mendel și-a efectuat experimentele și a formulat modele de importanță universală și fundamentală pentru dezvoltarea geneticii și a biologiei în general, cunoștințele științifice încă nu erau suficiente pentru a înțelege mecanismele moștenirii. De aceea, timp de mulți ani, opera lui Mendel a fost nerevendicată. Cu toate acestea, până la începutul secolului al XX-lea. situaţia în biologie s-a schimbat radical.
Au fost descoperite mitoza și meioza, legile lui Mendel au fost redescoperite. Independent unul de celălalt, cercetătorii din Germania și Statele Unite au propus ca factorii ereditari să fie localizați pe cromozomi. În 1906, R. Pennet a descris pentru prima dată încălcarea legii mendeliane a moștenirii independente a două trăsături. Când a pus în scenă o încrucișare dihibridă clasică de plante de mazăre dulce care diferă prin culoarea florii și forma polenului, Pennet nu a obținut împărțirea așteptată de 9:3:3:1 în a doua generație. Hibrizii F2 au avut doar fenotipuri parentale într-un raport de 3:1, adică nu a avut loc nicio redistribuire a trăsăturilor.
Treptat, s-au acumulat din ce în ce mai multe astfel de excepții, care nu respectau legea moștenirii independente. A apărut întrebarea, cum sunt exact genele localizate pe cromozomi? La urma urmei, numărul de trăsături și, prin urmare, numărul de gene din fiecare organism este mult mai mare decât numărul de cromozomi. Aceasta înseamnă că fiecare cromozom conține multe gene responsabile pentru diferite trăsături. Cum se moștenesc genele situate pe același cromozom?
■ Lucrarea lui T. Morgan. La aceste întrebări au răspuns un grup de oameni de știință americani condus de Thomas Hunt Morgan (1866-1945). Lucrând la un obiect genetic foarte convenabil - musca de fructe Drosophila, au făcut o treabă grozavă în studierea moștenirii genelor.
Oamenii de știință au descoperit că genele situate pe același cromozom sunt moștenite împreună, adică sunt legate. Acest fenomen a fost numit legea morgan, sau legea moștenirii legate . Se numesc grupuri de gene situate pe același cromozom grup de ambreiaj. Deoarece genele identice sunt localizate în cromozomi omologi, numărul de grupuri de legătură este egal cu numărul de perechi de cromozomi, adică numărul haploid de cromozomi. O persoană are 23 de perechi de cromozomi și, prin urmare, 23 de grupuri de legătură, un câine are 39 de perechi de cromozomi și 39 de grupuri de legătură, un bob de mazăre are 7 perechi de cromozomi și 7 grupuri de legătură etc. Trebuie remarcat că Mendel a fost surprinzător de norocos. la stabilirea încrucișărilor di-hibride: genele responsabile pentru diferite trăsături (culoarea și forma mazărelor) au fost localizate pe diferiți cromozomi. Ar fi putut fi altfel, iar atunci modelul divizării independente nu ar fi fost descoperit.
Rezultatul muncii grupului lui T. Morgan a fost crearea în 1911 a teoria cromozomiala a ereditatii.
Luați în considerare principalele prevederi ale teoriei cromozomiale moderne a eredității.
Unitatea de ereditate este o genă, care este un segment al unui cromozom.
Genele sunt localizate în cromozomi în locuri strict definite (locuși), iar genele alelice (responsabile pentru dezvoltarea unei trăsături) sunt localizate în aceleași loci ai cromozomilor omologi.
Genele sunt localizate pe cromozomi într-o ordine liniară, adică una după alta.
■Defecțiunea ambreiajului. Cu toate acestea, în unele încrucișări, la analiza moștenirii genelor situate pe același cromozom, a fost găsită o tulburare de legătură. S-a dovedit că uneori cromozomi omologi perechi pot face schimb de regiuni omoloage identice între ele. Pentru ca acest lucru să se întâmple, cromozomii trebuie să fie amplasați în imediata apropiere unul de celălalt. Această abordare temporară în perechi a cromozomilor omologi se numește conjugare. În acest caz, cromozomii pot face schimb de loci situati unul vizavi de celălalt care conțin aceleași gene. Acest fenomen se numește crossing over.
Amintiți-vă diviziunea meiozei, în timpul căreia se formează celulele germinale. În profaza primei diviziuni meiotice, în timpul formării unei bivalente (tetrade), când cromozomii omologi dublați stau paralel unul cu celălalt, poate avea loc un schimb similar. Un astfel de eveniment duce la o recombinare a materialului genetic, crește diversitatea descendenților, adică crește variabilitatea ereditară și, prin urmare, joacă un rol important în evoluție.
■ Hărți genetice. Fenomenul schimbului de gene alelice între cromozomii omologi a ajutat oamenii de știință să determine locația fiecărei gene pe cromozom, adică să construiască hărți genetice. Harta genetică a unui cromozom este o diagramă a aranjamentului reciproc al genelor situate pe același cromozom, adică în același grup de legătură. Construirea unor astfel de hărți prezintă un mare interes atât pentru cercetarea fundamentală, cât și pentru rezolvarea unei varietăți de probleme practice. De exemplu, hărțile genetice ale cromozomilor umani sunt foarte importante pentru diagnosticarea unui număr de boli ereditare severe.
În prezent, hărțile genetice simple sunt înlocuite cu hărți genetice moleculare, care conțin informații despre secvențele de nucleotide ale genelor.
Întrebări pentru autocontrol
1. Ce este moștenirea legată?
2. Ce sunt grupurile de legare a genelor?
3. Care este cauza tulburării de legătură genică?
4. Care este semnificația biologică a schimbului de gene alelice între cromozomii omologi?
5. Teoria moștenirii legate este confirmată citologic?
Potrivit lui G. Mendel, genele sunt moștenite independent și sunt localizate pe diferiți cromozomi. Diheterozigotul AaBb sub forma unei formule citologice poate fi reprezentat după cum urmează:
În timpul meiozei, formează 4 tipuri de gameți: AB, Ab, aB, ab.
În 1911–1912 Geneticistul american T. Morgan a demonstrat că genele situate pe același cromozom pot fi moștenite legate (împreună). Acest lucru se datorează faptului că există mult mai multe gene decât cromozomi. Co-moștenirea genelor limitându-și combinația liberă, T. Morgan a sunat legarea genelor sau moștenirea legată. În cazul în care genele sunt moștenite legate, genotipul diheterozigotului AaBb folosind formula citologică poate fi reprezentat după cum urmează:
În timpul meiozei, dacă există o legătură completă a genelor, formează 2 tipuri de gameți - AB, ab. Astfel de gameți sunt numiți non-încrucișați.
Cercetările efectuate de T. Morgan și studenții săi au arătat că genele sunt schimbate în mod regulat într-o pereche omoloagă de cromozomi. Procesul de schimb de secțiuni identice de cromozomi omologi cu genele localizate în ele se numește decusatie cromozomală sau trecere peste. Crossing over oferă noi combinații de gene situate pe cromozomi omologi . Prin urmare, din cauza fenomenului de crossing over, pe lângă non-crossover, există și gameți încrucișați - Ab, aB.
Legea lui T. Morgan este formulat astfel: genele situate pe același cromozom sunt moștenite legate (împreună), iar puterea legăturii depinde de distanța dintre gene.
Principalele prevederi ale teoriei cromozomilor lui T. Morgan:
1) genele sunt localizate pe cromozomi, iar cromozomi diferiți conțin un număr inegal de gene. Setul de gene din fiecare cromozomi neomologi este unic;
2) genele sunt localizate pe cromozom într-o anumită secvență liniară;
3) genele formează grupuri de legătură și pot fi moștenite legate. Numărul de grupuri de legătură corespunde setului haploid de cromozomi (o persoană are 23 de grupuri de legătură);
4) între cromozomi omologi este posibilă o încrucișare (încrucișare) și un schimb de gene alelice, iar încrucișarea are loc atunci când există o distanță între gene;
5) distanța dintre gene din cromozom este proporțională cu frecvența de încrucișare și este exprimată ca procent de încrucișare.
Procentul de încrucișare între gene este calculat prin formula:
X= ––––
a este numărul de indivizi încrucișați dintr-o clasă; b este numărul de indivizi încrucișați din altă clasă;
n este numărul total de indivizi; Х – % de încrucișare între gene (distanța dintre gene în % de încrucișare).
Distanța dintre gene se măsoară în unități speciale - morganides. 1 morganidă (1 M) = 1% crossing over.
În studiile genetice, a fost folosită o muscă de fructe, care are un cariotip format din 8 cromozomi, are fertilitate și multe trăsături alternative. T. Morgan a luat în considerare situaţia când legarea completă a genelor și încălcarea legăturii genelor (încrucișare) apar la un individ în același timp. În prima etapă, indivizi homozigoți (P ♀AABB x ♂aabb) au fost prelevați pentru încrucișare. Ca rezultat, în prima generație, toți indivizii erau diheterozigoți (AaBb).
Trăsătură genetică
O culoare gri a corpului
și culoarea întunecată a corpului
În aripi lungi
b aripi rudimentare.
Legătura completă a genelor
P ♀ ══x ♂ ══
G AB, ab ab - gameti masculini
(gameți feminini non-încrucișați)
F1 ══; ══
Cu o legătură completă a genelor, se observă o divizare 1:1. În total, s-au obținut 83% dintre indivizii care au moștenit fenotipul părinților, deoarece aceștia s-au format din gameți neîncrucișați. Dintre aceștia, indivizii cu corp gri, aripi normale au reprezentat 41,5% și indivizii cu corp negru, aripile rudimentare au reprezentat, de asemenea, 41,5%.
Tulburare de legătură genică
P ♀ ══ x ♂ ══
G Ab, aB ab - gameti masculini
(Gameți feminini încrucișați)
F1 ══; ══
Cu o legătură incompletă a genelor, se observă, de asemenea, divizarea 1:1. În total, s-au obținut 17% dintre indivizii cu fenotipuri recombinate, deoarece s-au format din gameți încrucișați. Sunt chemați indivizii cu o nouă combinație de trăsături recombinante. Dintre recombinanți, muștele cu corp gri, aripile rudimentare au reprezentat 8,5%, iar cu corp negru, aripile normale au reprezentat și 8,5%.
Întrebarea 1. Ce este moștenirea legată?
Moștenirea legată este moștenirea comună a genelor situate pe același cromozom (adică într-o moleculă de ADN). De exemplu, la mazărea dulce, genele care determină culoarea florilor și forma polenului sunt aranjate în acest fel. Ele sunt moștenite într-un mod legat, prin urmare, la încrucișarea hibrizilor din a doua generație, fenotipurile parentale se formează într-un raport de 3: 1, iar divizarea 9: 3: 3: 1, caracteristică încrucișării dihibride cu moștenire independentă, nu nu apare.
Întrebarea 2. Ce sunt grupurile de legare a genelor?
Un grup de legătură este o colecție de gene situate pe același cromozom. Deoarece cromozomii omologi conțin aceleași gene, numărul de grupuri de legătură este egal cu numărul haploid de cromozomi (23 la om, 7 la mazăre, 4 la Drosophila).
Întrebarea 3. Care este cauza tulburării de legătură genică?
Motivul pentru întreruperea legăturii genelor este schimbul de secțiuni de cromozomi omologi în profaza I a diviziunii meiotice. Amintiți-vă că în acest stadiu, cromozomii perechi sunt conjugați, formând așa-numitele benzi bivalente. Formarea bivalenților poate duce la încrucișarea cromozomilor, ceea ce creează posibilitatea schimbului de regiuni ADN omoloage. Dacă se întâmplă acest lucru, atunci grupurile de legătură își schimbă conținutul (conțin alte alele ale acelorași gene) și pot apărea la descendenți indivizi cu un fenotip care diferă de cel părinte.
Întrebarea 4. Care este semnificația biologică a schimbului de gene alelice între cromozomii omologi?material de pe site
Încrucișarea cromozomilor (în rest - crossing over) duce la recombinarea materialului genetic și formarea de noi combinații de alele genelor din grupul de legătură. În același timp, diversitatea descendenților crește, adică crește variabilitatea ereditară, ceea ce are o mare importanță evolutivă. Într-adevăr, dacă, de exemplu, la Drosophila, genele care determină culoarea corpului și lungimea aripilor se află pe același cromozom, atunci, încrucișând linii pure de muște cenușii cu aripi normale și muște negre cu aripi scurtate, în absența crossing over, nu vom obține niciodată alte fenotipuri. Existența unei încrucișări de cromozomi permite apariția (în câteva procente din cazuri) a muștelor cenușii cu aripi scurte și a muștelor negre cu aripi normale.
Întrebarea 5. A fost confirmată citologic teoria moștenirii legate?
Teoria moștenirii legate de Thomas Hunt Morgan (1866-1945) este confirmată de observațiile citologice. S-a demonstrat că în timpul diviziunii, cromozomii diverg complet către diferiți poli ai celulei. În consecință, genele situate pe același cromozom în timpul meiozei cad într-un singur gamet, adică sunt moștenite într-adevăr într-un mod legat.
Nu ați găsit ceea ce căutați? Utilizați căutarea